 Revista Científica Revista Científica
Vol. 17. No. 29, p. 78-86 / noviembre 2017 |
ISSN 1998-8850
URL: http://lacalera.una.edu.ni
DOI: https://doi.org/10.5377/calera.v17i29.6528
correo: lacalera@ci.una.edu.ni
|
| recursos naturales |
Diversidad y distribución de la macrofauna edáfica en dos sistemas de manejo de Moringa oleifera
(Lam.): relación con las propiedades del suelo
Diversity and distribution of edaphic macrofauna in two management systems of Moringa oleifera
(Lam.): Relationship with soil properties
|
|
Alvaro Noguera-Talavera1, Nadir Reyes-Sánchez2, Bryan Mendieta-Araica2
1Docente Investigador, Facultad de Recursos Naturales y Ambiente / 2 Docentes Investigadores, Facultad de Ciencia Animal. Para
correspondencia: Alvaro.noguera@ci.una.edu.ni
|
 |
RESUMEN
Con el objetivo de determinar la relación entre diversidad y distribución
vertical de la macrofauna edáfi ca, y propiedades físico-químicas
del suelo, se llevó a cabo un estudio en predios de la Universidad
Nacional Agraria. Los ensayos tuvieron una duración de 9
meses, en los que se implementaron prácticas de manejo agroecológico,
y manejo convencional en ocho unidades experimentales con
dimensiones de 15 m x 30 m, seleccionadas de manera aleatoria.
La metodología de muestreo fue la desarrollada por el Programa
Internacional Biología y Fertilidad del Suelo Tropical. Fueron levantados
12 monolitos de 25 cm x 25 cm por práctica de manejo,
a una profundidad de 30 centímetros. Se realizó identifi cación taxonómica
a nivel de Phylum, clase, orden y familia; así como de
grupos funcionales detritívoros, ingenieros del suelo, herbívoros y
depredadores. Fue registrada mayor densidad de macrofauna en el
sistema de conversión agroecológica (7424 ind/m2) en comparación
al sistema de manejo convencional (1984 ind/m2), con alta concentración
de individuos (7488 ind/m2) en los primeros 20 centímetros
del perfi l; y densidad proporcional del 56.2% en la profundidad 0
a 10 centímetros. Taxonómicamente se determinó dominancia de
la clase Insecta en ambos sistemas de manejo. La distribución de
familias de la macrofauna edáfi ca fue diferente entre sistemas, con
evidente influencia del rol por profundidad de muestreo. El grupo
funcional ingenieros del suelo predominó en conversión agroecológica;
mientras los detritívoros lo hicieron en manejo convencional.
Fue determinada la influencia de la calidad de micro hábitat en las
propiedades físico-químicas del suelo con el componente biológico;
demostrándose efecto de las prácticas de manejo sobre la densidad,
diversidad y distribución de la macrofauna edáfica
Palabras Clave: Macrofauna edáfica, Conversión agroecológicas,
Funcionalidad, Propiedades de suelo
|
ABSTRACT
With the aim to determine the relation between diversity and distribution
of edaphic macrofauna and soil properties, a study was
performed at National Agrarian University farm. International Program
for Biology and Fertility of Tropical Soil sampling methodology
was used. A taxonomical classification was done at phylum,
class, order and family level as well functional groups such as detritivores,
soil engineers, herbivores and predators. A higest density
of macrofauna was registered on agroecological consersion system
(7424 ind/m2) in comparision with conventional management system
(1984 ind/m2), with major individual concentration (7488 ind/
m2) on the fi rst 20 centimeters of the soil; and population density
of 56.2% in 0 to 10 centimeters strata. The number of taxas show
dominance of Insecta at both management systems. About family
taxa, the edaphic macrofauna distrubution was diferent between
systems with influence of the funtionality in each soil strata. The
soil engenier group was the dominan on agroecological consersion
system; while detritivorous group was dominan in conventional
system. Was determinate the influence of hábitat over physic and
chemical soil properties, and the biological populations; founding
efecto of management over the density, diversity and distribution of
the edaphic macrofauna.
Keywords: Edaphic macrofauna, Agroecological conversion, Functionality,
Soil properties
|
|
L
a agricultura convencional ha sido contextualizada
en diversos estudios (Altieri y Nicholls, 2012; Nieto
et al., 2013; Altieri et al., 2015, Hatt et al., 2016)
como factor causante de la actual crisis ambiental,
que provoca baja sostenibilidad de la agricultura (Gliessman,
2013), degradación del suelo, pérdida de biodiversidad por
simplificación de los ecosistemas (Hatt et al., 2016; Altieri
et al., 2017), y alteración del ciclo hidrológico, además de la
contaminación de aguas subterráneas y superficiales (Durán
y Suárez, 2013).
Mantener la calidad de los suelos y la productividad
de los cultivos es un desafío importante para la agricultura
moderna. Prácticas de manejo agroecológico que tienden a
incrementar el contenido de carbono del suelo y a la vez favorecer
la supervivencia y actividad microbiana pueden contribuir
a la sustentabilidad de los sistemas agrícolas (Ferreras
et al., 2015); efectos que pueden ser evaluados a través de la
composición y funcionalidad de las propiedades biológicas
del suelo, entre la que destaca la macrofauna, por la facilidad
de colectarla.
La macrofauna edáfica agrupa a los invertebrados
mayores de 2 mm de diámetro (Cabrera, 2012), los que
presentan características eco fisiológicas que permiten ser
utilizada como indicadores de monitoreo y diagnóstico de
intensidad de uso del suelo (Díaz et al., 2014), estado de conservación
o perturbación y, salud (Cabrera, 2012; Vieira Da
Cunha, 2012); así como del efecto en el tiempo de prácticas
productivas (Soccorras e Izquierdo, 2014; Gómez et al.,
2016).
Como elemento técnico con enfoque agroecológico
en la promoción de Moringa oleifera Lam., como especie
con múltiples usos, a nivel de sistemas productivos intensivos,
en contraposición a los sistemas con alta demanda de
insumos; se identificó la necesidad de estudiar las ventajas
que implica que en sistemas de cultivos diversificados y con
bajos insumos; se pueden obtener servicios asociados a la
recuperación y salud del suelo por medio del incremento y
manejo de la diversidad funcional de la macrofauna edáfica.
Fue en el contexto de la utilización de bioindicadores del
efecto de prácticas de manejo en sistemas productivos de
Moringa oleifera Lam., que se planteó como objetivo analizar
la relación entre diversidad, distribución de la macrofauna,
y propiedades físico-químicas del suelo.
MATERIALES Y MÉTODOS
Características edafológicas y climatológicas de la zona,
o lugar de los experimentos. El trabajo fue realizado entre
Junio del 2013 y Marzo del 2014, en la unidad experimental
Santa Rosa de la Universidad Nacional Agraria, localizada
geográficamente en Managua, en las coordenadas
12o09’30.65”N, 86o10’06.32”W (Mendieta-Araica, 2011), a
una altitud de 50 msnm.
La precipitación y temperatura media anual históricas
registradas son 1099 mm y 27o C respectivamente, y
humedad relativa de 74% (INETER, 2015); predominando
dos estaciones definidas por una época seca que va desde
Noviembre a Abril, y una época lluviosa de Mayo a Octubre
En el año 2013 la precipitación anual en el área fue
de 1070.4 mm, menor en comparación a la media histórica,
siendo Junio (285.3 mm) y Septiembre (330.2 mm), (figura
1) los de mayor precipitación (INETER, 2015).
El suelo pertenece al orden taxonómico Andisol,
y clase textural Franco Arcilloso –Arenoso; y buen drenaje
(Mendieta-Araica, 2011).
Procedimiento experimental
Duración de los ensayos. Los ensayos tuvieron una duración
de nueve meses, con fecha de establecimiento junio 2013, y
cosecha en marzo 2014, tiempo durante el cual se aplicaron
prácticas de manejo agroecológico y de agricultura convencional
según la planificación y enfoque de cada sistema.
El área con manejo agroecológico correspondió a
un lote de 1 hectárea, y el agroecosistema con manejo convencional
consistió en un área de plantación, con una extensión
de 5 hectáreas; utilizándose, en ambos casos, un área
efectiva de muestreo de 0.18 hectáreas. En cada sistema fueron
delimitadas cuatro unidades de muestreo de forma rectangular
(15m x 30m). El método definido para muestrear la
macrofauna edáfica fue sistemático con monolitos separados
15 m entre sí, distribuidos en transecto diagonal dentro de las
unidades de muestreo.
Vegetación colindante a los sistemas de manejo
Conversión agroecológica. La biota complementaria al agrosistema
estuvo compuesta por cercas vivas con especies arbóreas
como Eucalyptus camaldulensis (Dehnh.), Azadirachta
indica (A. Juss), Cordia dentata (Poir.), Pithecelobim
dulce (Roxb.) Benth.), Albizia saman (Jacq.) Muell.), y Stemmadenia
obovata (Hook. & Arn.) K. Shum.); mientras los
lotes circundantes corresponden a uso de suelo agrícola con
Moringa oleifera (Lam.) y Sorghum bicolor (L.).
Manejo convencional. La biota complementaria al agrosistema
estuvo constituidas por cercas vivas con especies arbóreas
como Neem Azadirachta indica (A. Juss), Spondias sp,
y Albizia saman (Jacq.) Muell.); mientras los lotes circundantes
corresponden a producción de pasturas como CT-15
y Cynodon nlemfuensis (Vanderryst.); además de cultivos
como Saccharum officinarum (L.), Moringa oleifera (Lam.)
y Sorghum bicolor (L.).
Diseño experimental.
El diseño consistió en unidades experimentales
de forma rectangular, seleccionadas de manera
aleatoria (DCA). Cada unidad con plantas establecidas a distanciamiento
3m x 3m, en arreglo lineal.
Tratamientos.
Tratamiento 1: Manejo convencional, el que
consistió en área de monocultivo de moringa, con actividades
de preparación mecanizada del suelo, control mecanizado y
químico de arvenses; fertilización inorgánica, y riego. El lote
que correspondió a este tratamiento tiene una historia de uso
de pastura permanente de pasto estrella (Cynodon nlemfuensis).
Tratamiento 2: Conversión agroecológica, cuyo enfoque
fue el establecimiento de un sistema de policultivo a partir
del sistema convencional, labranza mínima en la preparación
del suelo, rotación de leguminosas, fertilización orgánica con
compost, incorporación de abonos verdes, sin riego y, control
de arvenses con cobertura de leguminosas. El lote que
correspondió a este tratamiento tiene una historia de uso de
barbecho forestal o tacotal.
Evaluaciones y mediciones.
Se realizó colectas de macrofauna
edáfica antes de la cosecha o explotación de los sistemas.
El periodo de colecta fue entre diciembre 2013 y enero
del 2014, al finalizar la época lluviosa; definido a partir de los
criterios propuestos por Cabrera et al., (2011).
Para la colecta se utilizó la metodología propuesta
por el Programa Internacional “Biología y Fertilidad del
Suelo Tropical” (Lavelle et al., 2003), y validada por Zerbino,
(2010). De cada sistema de manejo se extrajeron 12
monolitos de suelo de 25 x 25 cm, hasta una profundidad de
30 centímetros; siendo los estratos: Hojarasca, 0 a 10 cm, 10
a 20 cm y 20 a 30 cm.
Los especímenes colectados fueron depositados en
viales con alcohol al 70% y luego identificados a nivel de clase,
y familia por profundidad de muestreo, mediante el uso de
claves desarrolladas por Roldan, (1988); Castner, (2000); y
Marshall, (2008), en el laboratorio de Biología de la Facultad
de Recursos Naturales y del Ambiente.
La macrofauna fue clasificada en detritívoros, herbívoros,
ingenieros del suelo y depredadores según los grupos
funcionales propuestos por Cabrera et al., (2011). Los
resultados se basan en la densidad (individuos por m2) de
cada taxón, y grupo funcional identificado; y diversidad por
sistema de manejo y profundidad de muestreo; mientras la
diversidad, y composición a nivel de familia fue analizada
como indicador relacionado a la salud del suelo según Rendón
et al., (2011) y, la interacción con el cultivo moringa.
|
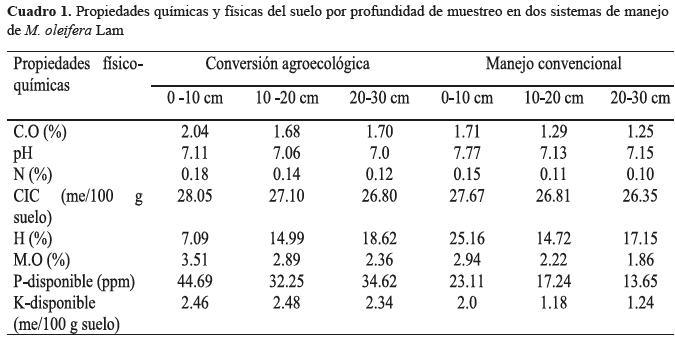
pH: Relación agua-suelo 1:2.5, Hester, (1930) en A.O.A.C, (1980). M.O: Combustión húmeda de Walkey y
Black, (1934), en A.O.A.C, (1980). N: Kjendalh, (1883) en A.O.A.C, (1980). P: OLSEN, (1954) en A.O.A.C,
(1980). Humedad%: Método gravimétrico en A.O.A.C, 1980). C.O: Combustión húmeda de Walkey y Black,
(1934), en A.O.A.C, (1980).
Del conjunto de monolitos se colectó dos muestras
compuestas de 2 kilogramos de suelo, para determinar propiedades
químicas y físicas en el Laboratorio de Suelos y
Agua (LABSA) de la Universidad Nacional Agraria (Cuadro
1).
|
|
Procesamiento estadístico.
La estadística no paramétrica
(Prueba de Kruskal-Wallis) fue empleada para determinar
diferencias en la variación de la densidad por taxón, grupo
funcional por sistema de manejo y profundidad de muestreo.
Como parte de la diversidad de la macrofauna, a nivel de
sistema de manejo para los taxones clases y ordenes, se determinó
el índice de dominancia de la comunidad (D) mediante
el método propuesto por Turner y Garner, (1991) con
comparaciones de “t” student en el programa PAST versión
1.29 (Hammer y Harper, 2004).
Un análisis de conglomerados para determinar la
probabilidad de similitud de familias por sistema de manejo
y profundidad de muestreo fue realizado determinándose el
índice de Jaccard a partir del método propuesto en Moreno,
(2001). Este índice expresa el grado en que dos o más muestras
son semejantes en los niveles taxonómicos presentes en
ellas, utilizándose en este trabajo para el nivel de familia.
Análisis de componentes principales (ACP) fue realizado
para determinar la asociación entre la diversidad y densidad
de la macrofauna con las propiedades físico-química del suelo.
|
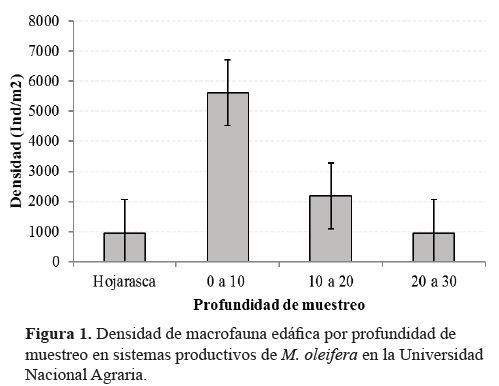
RESULTADOS Y DISCUSIÓN
Densidad de organismos de la macrofauna edáfica por
profundidad y sistemas de manejo. Fue registrada una
densidad total de 9408 individuos de la macrofauna edáfica.
La mayor densidad
acumulada de organismos
fue registrada en los
primeros 20 centímetros
del suelo (7488 ind/m2);
con alta concentración en
la profundidad 0 a 10 cm
(5296 ind/m2); y densidad
similar entre hojarasca
con la profundidad 20
a 30 centímetros (figura
1).
Diferencia
estadísticamente significativa
(p<0.05) fue registrada
en la densidad
de organismos entre la
profundidad 0 a 10 cm
respecto a las demás.
|
 |
|
Aratat et al., (2002), reportaron como
patrón en la distribución de la macrofauna del
suelo, alta densidad de organismos en los primeros
10 centímetros de profundidad, con acumulación
hasta de un 80.2% en relación a la densidad
total, proporción que en el actual estudio
correspondió a 56.2%, mientras Fernández et al.,
(2015) reportaron altas densidades en la profundidad
10 a 20 centímetros, y a diferencia de este
estudio, notable similitud en la densidad de macrofauna
edáfi ca, con la de la hojarasca.
Ararat et al., (2002) y Fernández et al.,
(2015), relacionaron la alta concentración de organismos
en los primeros centímetros del perfil
de suelo con la utilización de cultivo de cobertura
de leguminosas (en este caso canavalia, gandul,
caupi), que induce una disminución en la intensidad
de luz y del impacto de las gotas de lluvia;
además de la reducción en la temperatura
del suelo; e implica procesos biológicos de éste como son:
retención de humedad, aireación, mantenimiento de la biodiversidad
funcional y eficiencia del ciclo de nutrientes, provocada
según Duval et al., (2014) por el incremento gradual de
compuestos asociados a la fracción mineral, y por tanto una
fracción más transformada de carbono o materia orgánica lábil.
Padoa et al., (2012) determinaron que la alta concentración
de organismos de la macrofauna es producto que
este horizonte es una interface entre el liter y el dominio de
suelo funcional.
Por su parte, Vega et al., (2014), reportaron las mayores
densidades a nivel de la hojarasca, y al igual que en
este trabajo, notable reducción de la densidad en la profundidad
20 a 30 centímetros. En dicho trabajo, se identificó como
factores causales la concentración de biomasa de leguminosas
que implicó incremento en los recursos trófi cos, microhábitats
e interacciones funcionales.
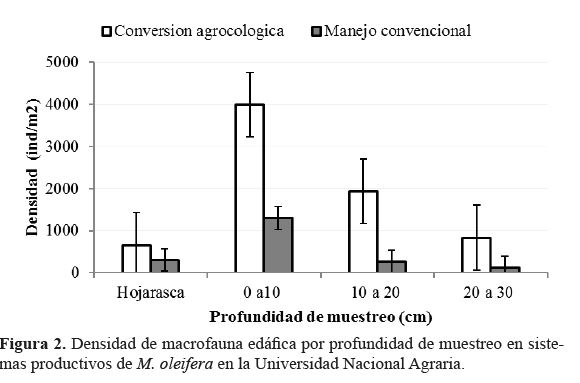
En el sistema conversión agroecológica fue registrada
mayor densidad de macrofauna edáfica (7424 ind/m2),en comparación a manejo convencional (1984 ind/m2), resultado
que coincide con el reportado por Matienzo et al.,
(2015), quienes lo atribuyen al grado de complejidad del sistema,
expresado en la diversificación y prácticas de manejo;
enfatizando, además en la importancia de los ambientes seminaturales
persistentes en la circundancia de los sistemas.
La densidad de macrofauna por sistema de manejo
mostró el mismo resultado de alta concentración en los
primeros 10 centímetros, mientras se determinó diferencias
estadísticas (p<0.05) en la densidad entre sistemas; así como
en la densidad de macrofauna en las profundidades 0 a 10 y
10 a 20 en ambos sistemas de manejo (p<0.05).
En el sistema bajo conversión agroecológica se registró
la mayor densidad en todas las profundidades de muestreo
(figura 2).
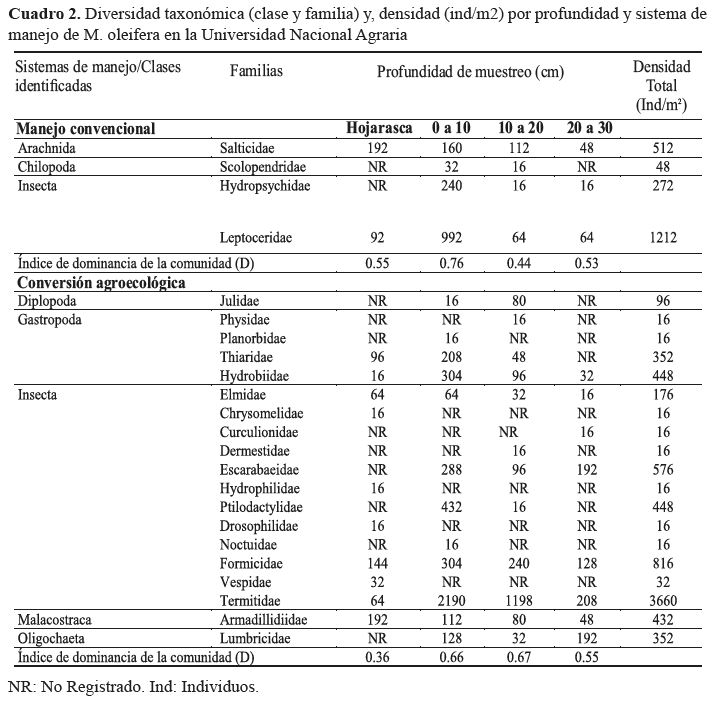
Desde el punto de vista taxonómico se identificó
asociación entre la profundidad y la riqueza de clases de la
macrofauna, registrándose igual número (8) en las profundidades 10 cm y 20 cm, lo que signifi có más diversidad, densidad
y mejor distribución de la macrofauna que la reportada a
nivel de hojarasca y 30 centímetros (5 clases).
|
 |
|
El mayor número de clases de la macrofauna fue
registrado en conversión agroecológica (5), en comparación
al sistema de manejo convencional (3); con dominancia por
alta densidad de la clase Insecta en ambos sistemas de manejo
(cuadro 2).
Los resultados relacionados a la profundidad de
muestreo confi rman la relación con la composición y densidad
de la macrofauna en ambos sistemas de manejo; registrándose
diferencias significativas (p<0.05).
A nivel de familia, las pertenecientes a la clase Insecta
reportaron valores altos de densidad en ambos sistemas
de manejo (cuadro 2). Leptoceridae fue la familia más
dominante en manejo convencional por la densidad que registró
(1212 ind/m2), mientras en conversión agroecológica
Termitidae (3660 ind/m2), Formicidae (816 ind/m2) y Escarabaeidae
(576 ind/m2) fueron las familias con mayor densidad
(cuadro 2).
|
 |
|
Familias no pertenecientes a la clase Insecta y que
registraron importante presencia en los sistemas fueron: Salticidae
(512 ind/m2) en manejo convencional, y Armadillidiidae
(432 ind/m2) y Lumbricidae (352 ind/m2) en conversión
agroecológica. La densidad registrada para Lumbricidae fue
inferior a reportes realizados por Ararat et al. (2002), Duran
y Suarez, (2013), y Fernández et al., (2015) determinándose
baja densidad de este grupo, que según Ararat et al., (2002)
tiene como funciones primordiales la formación de macroagregados,
transformación de sustancias orgánicas e inorgánicas,
mejoramiento de las tasas de infiltración dentro del perfil
del suelo, aireación, entre otras.
La presencia de clases por profundidad mostró una
notable equitatividad entre sistemas de manejo; con excepción
de Oligochaeta y Diplopoda en conversión agroecológica;
y Chilopoda, en manejo convencional, provocada la ausencia
de representantes de dichas clases por la especificidad
ambiental de distribución en el perfil del suelo (Masin et al.,
2015), carencia de hábitats aptos; ya que para Oligochaeta
la distribución está asociada a alto contenido de humedad
y bajo grado de compactación del suelo (Fernandez et al.,
2015; Masin et al., 2015). Para ambos sistemas de manejo, la
compactación del suelo fue asumida como factor limitante,
por la historia de uso que refiere principalmente ganadería.
La presencia de familias
por profundidad, al igual
que para el nivel taxonómico
clases, permiten identificar
atributos como adaptación,
especificidad de hábitat, y
su relación con la estructura
y procesos biológicos
de la calidad del suelo; por
ejemplo, Termitidae estuvo
presente desde la hojarasca
hasta los 30 centímetros de
profundidad, lo que indica
alto grado de adaptación a diferentes
ambientes y hábitos
alimenticios (Sermeño-Chicas,
2013) por ser especie
pionera, asociada a raíces y
tallos de M. oleífera durante
fases no adultas de la planta,
y formadores de colonias
sobre ramas de árboles en la
fase adulta del árbol (Palada
y Chang, 2003); y transformadores
de la estructura del
suelo a través de la formación
de bioestructuras (Da Cunha
y Brandao, 2000; Sermeño-
Chicas, 2013).
Formicidae presenta
alta adaptación a variedad
de ambientes (Castro et al.,
2008; Gutiérrez-Rodríguez,
2014), alta reproducción, formadores
de colonias, depredadores y transformadores de la
estructura del suelo al mover (traslocar) grandes cantidades
de detrito a estratos inferiores del suelo (Castro et al.,
2008), por lo que, de acuerdo con Fernández et al., (2015),
pueden estar presente en diferentes profundidades del perfil
del suelo.
Otras familias con presencia en todos los estratos
del suelo hasta los 30 centímetros fueron: Hydrobiida en
manejo convencional y, Elmidae y Armadillidiidae en conversión
agroecológica (cuadro 2), los que biológicamente
indican notable concentración de materia orgánica en descomposición.
Diversidad de la macrofauna en los sistemas estudiados.
Al comparar la diversidad de la macrofauna edáfica
entre sistemas de manejo, se registró menor dominancia de
familias en conversión agroecológica, en todas las profundidades.
En manejo convencional, la diversidad basada en
el índice de dominancia de la comunidad registró alto valor
(D=0.76) en la profundidad 0 a 10 cm debido a la densidad
de las familias Leptoceridae, Salticidae e Hydropsychidae;
cuantificando menor dominancia en la profundidad 0 a 20
cm, producto de densidades más equitativas en las familias
que se registró.
En conversión agroecológica, la dominancia estuvo
concentrada en las profundidades 0 a 10 cm (D=0.66) y 10
a 20 cm (D=0.67), estando este parámetro en función de la
densidad o dominancia de la familia Termitidae (cuadro 2).
La menor dominancia de familias por profundidad
de muestreo se dió en hojarasca en ambos sistemas de manejo;
con ventaja a favor de alta diversidad y funcionalidad
de organismos en el sistema en conversión agroecológica
(D=0.36), en comparación a manejo convencional (D=0.55).
Los resultados a nivel de densidad y diversidad,
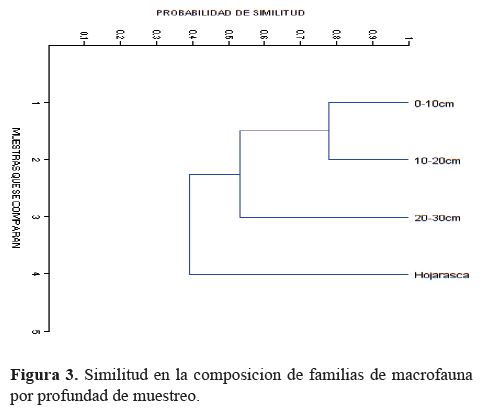
mostraron lo que se presenta en la figura 3, en cuanto a la
similitud de familias por profundidad, la que confirma un
ensamblaje de funciones en el perfil del suelo.
|
 |
|
La similitud de familias entre profundidades de
muestreo registró su valor más alto (J=0.78) en 0 a 10 y 10 a
20 cm, lo cual indica una notable igualdad en la composición
a nivel de este taxón entre ambas profundidades.
El valor de la similitud entre profundidades mostró
una reducción de 26% entre los primeros 20 cm del suelo,
en comparación con la profundidad 20 a 30 cm. Dicho indicador
registró la mayor reducción (40%) al comparar las
profundidades 0 a 30 cm con la composición de familias
registradas en la hojarasca. La reducción en la similitud de
familias entre profundidades fue asumida según lo expuesto
por Vega et al., (2014) quienes identificaron diferencias en
microhabitats, niveles de descomposición de la biomasa y,
por tanto, disponibilidad de materia orgánica, humedad, temperatura,
especialización de las especies a ciertos hábitats, y
procesos ecológicos propios de cada profundidad en el perfil
de suelo.
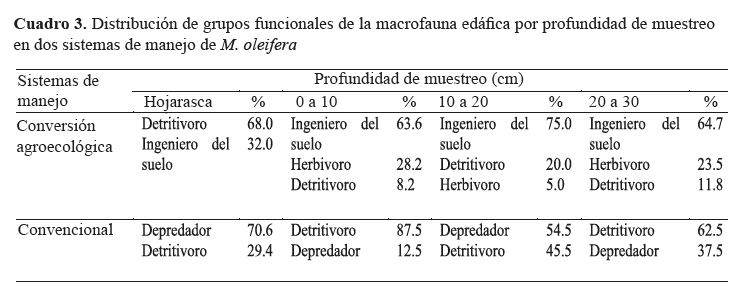
Grupos funcionales por profundidad.
Los grupos funcionales
de la macrofauna edáfica y su distribución en el perfil
del suelo, fueron elementos diferenciantes entre los sistemas de manejo, identificando mayor funcionalidad a favor de
cambios en la estructura y composición del suelo en el sistema
de conversión agroecológica, en el que se registró una
alta proporción de organismos clasificados como ingenieros
del suelo, que dominaron (proporción mayor al 50% de la
densidad total) en los 30 centímetros de profundidad de los
monolitos (cuadro 3).
Entre el grupo funcional ingenieros del suelo, sobresalieron
Termitidae y Formicidae con 48.3% y 10.7%
respectivamente, de la densidad total de organismos en conversión
agroecológica. Otro representante de este grupo funcional
como Lumbricidae (4.7%), registró valores bajos en
comparación al trabajo de Duran y Suarez, (2013), quienes
reportaron la dominancia de miembros de Lumbricidae en
una proporción de 78.5% de todos los individuos pertenecientes
al grupo funcional ingenieros del suelo.
En conversión agroecológica, la dominancia proporcional
de detritívoros (68.0%) en hojarasca estuvo relacionada
a la biomasa de cultivos de cobertura en la superficie
del suelo, cuyo propósito fue brindar cobertura para retención
de humedad, contribuir a la fertilidad, y controlar la
densidad de arvenses; demostrando con este resultado que la
incorporación de cobertura al suelo contribuye a incrementar
la presencia de grupos de organismos que aportan a los cambios
en las propiedades del suelo.
Las familias más representativas del grupo detritívoros
fueron: Hydrobiidae, Ptilodactylidae ambos con
6.03%, Armadillidiidae con 5.8% y Thiaridae con 4.7%. La
distribución de familias en el perfil del suelo fue diferente,
con la presencia de Ptilodactylidae únicamente de 0 hasta 20
cm, Thiaridae, con presencia tanto en la hojarasca hasta 20
cm, y Armadillidiidae, e Hydrobiidae en todo el perfil caracterizado
(cuadro 2).
La presencia de herbívoros desde 0 a 30 centímetros
estuvo caracterizada por un lado por la existencia en estado
larvario de organismos plaga de raíces, representantes de la
familia Escarabaeidae (Aragón et al., 2005), que con base en
lo reportado por Ararat et al. (2002) puede ser útil como indicador
de acumulación de detritos y materia orgánica en descomposición
por su condición de saprófito; asociado además,
a la implementación de labranza mínima en la preparación de
suelo, y manejo de cobertura de residuos de cosecha.
En el sistema de manejo convencional tanto en la
hojarasca, como en las profundidades hasta los 30 centímetros,
se registró dominio de los grupos detritívoros y depredadores
en una alternancia entre profundidades (cuadro 3).
En relación a los detritívoros, la familia dominante
fue Leptoceridae con 94% de la densidad de detritívoros y,
60.4% de la densidad total de organismos en dicho sistema de
manejo; mientras que el grupo funcional depredadores estuvo
representado únicamente por la familia Salticidae en una
proporción de 91% de la densidad del grupo y 23.3% de la
densidad total de macrofauna edáfica en el sistema.
|
 |
|
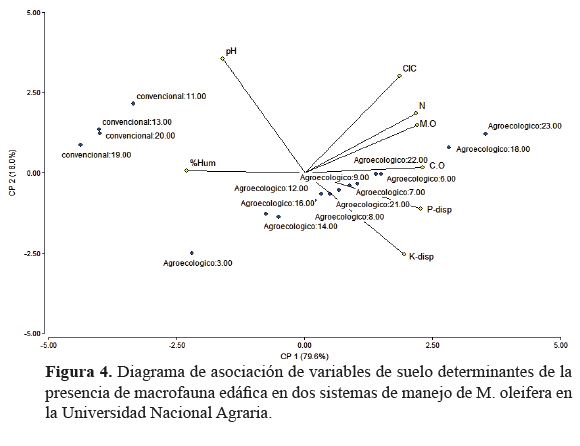
Asociación entre macrofauna edáfica y propiedades
del suelo. La determinación de asociación entre variables
edáficas y macrofauna del suelo a partir del análisis de
componentes principales presentó una alta confiabilidad al
explicar en los dos ejes de la componente principal 97.6% de
la variabilidad acumulada correspondiente a la densidad de
individuos por sistema de manejo (figuras 5 y 6).
El primer eje explicó el 79.6% de la varianza total
y separó a los sistemas, ubicando el sistema conversión
agroecológica a la derecha, en función de mayor asociación y
complementariedad entre las variables edáficas; y, por tanto,
mejor calidad de hábitat para la presencia, densidad y diversidad
de la macrofauna (figura 4). El segundo eje, ubicado en
la parte superior izquierda representó al sistema de manejo
convencional y explicó el 18% de la varianza total e identificó
el pH y el porcentaje de humedad con el hábitat asociado
a los grupos de macrofauna edáfica registrados.
Las variables edáficas relacionadas a la presencia
de macrofauna presentaron mayor asociación en conversión
agroecológica (figura 4). Las propiedades con mayor grado
de asociación entre sí fueron el porcentaje de Nitrógeno
y Materia orgánica, Capacidad de intercambio catiónico
y Carbono orgánico; sin embargo, las variables respuesta
asociadas a la presencia de macrofauna fueron contenido de carbono orgánico, fosforo
y potasio disponible, y
porcentaje de humedad, lo
que permitió determinar un
notable dinamismo en el ciclaje,
disponibilidad y flujo
de nutrientes en conversión
agroecológica.
|
 |
|
En manejo convencional
las variables respuesta
fueron el pH y el porcentaje
de humedad del suelo,
determinándose baja relación
entre sí. Los grupos de
mayor representación de la
macrofauna edáfica en este sistema tuvieron alta asociación
con el porcentaje de humedad del suelo.
Fue determinada asociación entre las familias más
representativas de la macrofauna y las variables edáficas,
siendo la excepción la familia Salticidae cuyo nicho está
circunscrito al estrato superficial del suelo, y Curculionidea;
explicando de esta manera la influencia del manejo de los
sistemas sobre grupos específicos y las características biológicas
del suelo.
La presencia de las familias Formicidae, Termitidae,
y Escarabaeidae, pertenecientes al grupo funcional ingenieros
del suelo, estuvo asociada a la disponibilidad de fosforo y
potasio.
Las familias Armadillidiidae, y Lumbricidae, también
incluidas dentro del grupo ingenieros del suelo presentaron
asociación con el porcentaje de humedad del suelo; al
igual que organismos de la familia Julidae, que ejercen el rol
de detritívoro, y cuya importancia radica en consumir hojas
de bajo valor nutritivo y excretarlas transformadas (fragmentadas),
facilitando el consumo para otros organismos de la
macrofauna edáfica (Chávez et al., 2016), incrementado la
materia en descomposición en el agroecosistema.
Padoa et al., (2012) determinó asociación
entre la abundancia biológica de la familia Julidae y
la relación Carbono/Nitrógeno en seis arreglos de cultivos
que tuvieron como prácticas la no labranza, el
uso de gandul como cobertura y rotaciones, prácticas
comunes al sistema de conversión agroecológica y
por tanto que refleja la relación entre las prácticas de
agricultura sostenible y la calidad biológica del suelo.
Para el caso de Lumbricidae, las variables
respuesta asociada a la presencia difiere del trabajo
realizado por Masin et al., (2015) en donde se determinó
asociación con la materia orgánica, capacidad
de intercambio catiónico, nitrógeno total y el pH; diferencia
dependiente del hábito epigeo o endógeno de
las especies de dicha familia. Padoa et al., (2012) por
su parte determinaron asociación entre la abundancia
y el nitrógeno contenido a los 10 centímetros de profundidad.
Castro et al., (2008) determinaron que especies de
la familia Formicidae al transportar restos vegetales y animales
hacia el interior del suelo concentran en sus nidos, y
alrededor altos niveles de fósforo lo que permite explicar la
asociación entre este elemento y la densidad de hormigas
cuantificadas en el sistema conversión agroecológica.
Complementariamente, Castro et al., (2008) encontraron
relación entre la distribución de Formicidae y la
humedad, comprobando que bajos valores de humedad incrementan
la densidad y diversidad; lo que permite adicionar otro elemento a la explicación de la presencia de Formicidae
en todo el perfil muestreado, debido a que el bajo porcentaje
de humedad registrada (Cuadro 1), el que fue menor al 30%
durante el periodo de evaluación generó un hábitat propicio
para este grupo.
|
 |
|
Fernández et al., (2015) evidenció la asociación
existente entre la variable humedad y pH con la riqueza y
densidad de especies de la macrofauna edáfica, por lo que se
puede explicar a través de esto la presencia de las familias
Scolopendridae y Leptoceridae las que presentaron asociación
con el porcentaje de humedad del suelo; por
el contrario, la presencia de Hydropsychidae estuvo
asociada a la variable pH.
|
CONCLUSIONES
Las prácticas asociadas al manejo de los sistemas
productivos de M. oleifera tuvieron efecto sobre
la distribución y diversidad de la macrofauna edáfica.
Se determinó mayor complementariedad
entre la distribución y diversidad de la macrofauna
edáfica y las variables químicas en el sistema
de conversión agroecológica.
Existe reciprocidad entre el tipo de práctica
relativa al manejo del cultivo de moringa
con las propiedades físico-químicas del suelo y;
por tanto, influencia en la densidad y funcionalidad
de los organismos de la macrofauna edáfica.
|
REFERENCIAS BIBLIOGRÁFICAS
Altieri, MA; Nicholls, CI. 2012. Agroecology scaling up for food sovereignty and resiliency. Sustainable Agriculture Reviews 11, 1
DOI 10.1007/978-94-007-5449-2_1.
Altieri, MA; Nicholls, CI; Henao, A; Lana, MA. 2015. Agroecology and design of climate change-resilient farming systems. Agronomy
Sustainable Development. DOI 10.1007/s13593-015-0285-2.
Altieri, MA; Nicholls, CI; Montalba, R. 2017. Technological approaches to sustainable agricultura at a crossroads: An agroecological
perspective. Sustainability. 9:(349).
Ararat, MC; Aristizábal, A; Prager-Mosquera, M. 2002. Efecto de cinco manejos agroecológicos de un Andisol (Typic Dystrandept)
sobre la macrofauna en el municipio de Piendamó, departamento del Cauca, Colombia. Acta Agronómica. 51(34):121-129.
Aragón, GA; Morón, MA; López-Olguín, JF; Cervantes-Peredo, LM. 2005. Ciclo de vida y conducta de adultos de cinco especies de
Phyllophaga Harris, 1927 (Coleoptera: Melolonthidae; Melolonthinae). Acta zoológica Mexicana. 21(2):87-99.
Cabrera, G.; Robaina, N; Ponce De León, D. 2011. Composición funcional de la macrofauna edáfica en cuatro usos de la tierra en las
provincias de Artemisa y Mayabeque, Cuba. Pastos y forrajes. 34(3):331-346
Cabrera, G. 2012. La macrofauna edáfica como indicador biológico del estado de conservación/perturbación del suelo. Resultados
obtenidos en Cuba. Pastos y forrajes. 35(4):349-363
Chávez, SL; Labrada, HY; Álvarez, FA. 2016. Macrofauna del suelo en ecosistemas ganaderos de montaña en Guisa, Granma, Cuba.
Comunicación corta. Pastos y forrajes. 39(3):111-115.
Castro, DS; Vergara, CC, Arellano, UC. 2008. Distribución de la riqueza, composición taxonómica y grupos funcionales de hormigas
del suelo a lo largo de un gradiente altitudinal en el Refugio de Vida Silvestre Laquipampa, Lambayeque-Perú. Ecología Aplicada.
7(12):90-103
Castner, J. 2000. Photographic Atlas of Entomology and Guide to Insect Identification. Feline Press. Gainesville, Florida. 174 pp.
Da Cunha, HF; Brandao, D. 2000. Invertebrates Associated with the Neotropical Termite Constrictotermes cyphergaster (Isoptera:
Termitidae, Nasutitermitinae). Sociobiology. 37:593-598.
Díaz, PM; Rionda, HM; Duhour, EA; Momo, RF. 2014. Artrópodos del suelo: Relaciones entre la composición faunística y la intensificación
agropecuaria. Ecología Austral. 24:327-334.
Durán, BE; Suárez, SJ. 2013. Fauna del suelo y hojarasca en arreglos agroforestales de la Amazonia Colombiana. Momentos en Ciencia.
10(1):59-66.
Duval, ME; De Sa Pereira, E; Iglesias, JO; Calantini, JA. 2014. Efecto de uso y manejo del suelo sobre las fracciones del carbono
orgánico en un Arguidol. Ciencia del suelo. 32(1):105-115.
Fernández, TM; Castellanos, GL; Fuentes, GM; Cairo, CP; Rajadel, AN; De Melo, PR. 2015. Macrofauna del suelo en cuatro fincas
en conversión hacia una producción agroecológica en el Municipio, Cruces, Cuba. Centro Agrícola. 42(1): 43-52.
Ferreras, L; Toresani, S; Bonel, B; Fernández, E; Bacigaluppo, S; Faggioli, V. 2009. Parámetros químicos y biológicos como indicadores
de calidad del suelo en diferentes manejos. Ciencia del Suelo. 27(1):103-114.
Gliessman, SR. 2013. Agroecología: Plantando las raíces de la resistencia. Agroecología. 8:19-26.
Gómez, PF; Godoy, MC; Coronel, JM. 2016. Macrofauna edáfica en ecosistemas y agroecosistemas de la Region Esteros del Iberá
(Corrientes, Argentina). Ciencia del suelo. 34(1):43-56.
Hammer, O; Harper, D.A.T. 2004. Paleontoloic Statistic (PAST versión 1.29). http://folk.uio.no/ohammer/past
Hatt, S; Artru, S; Brédart, D; Lassois, L; Francis, F; Haubruge, E. 2016. Towars sustainable food systems: the concept of agroecology
and how it questions current research practices. A review.Biotechnology, Agronomy, Sociaty and Environment. 20(S1):215-224.
Instituto Nicaragüense de Estudios Territoriales. (INETER). 2015. Boletín climático. Managua, Nicaragua. Dirección General de
Meteorología.
Lavelle, P; Senapati, P; Barros, E. 2003. Soil macrofauna. In: Trees, crops and soil fertility. Concepts and research methods. (Eds. G.
Schroth & F.L.Sinclair). CABI Publishing. UK. p. 303.
Marshall, S. 2008. 500 insects: a visual reference. Firefly Books Ltd. Ellicott Station Buffalo, New York. USA. 528 pp.
Masin, EC; Rodríguez, RA; Maitre, IM; Cerana, J; Hernandez, PJ; Anglada, M; Elizalde, HJ; Lallana, MC. 2015. Riqueza de lombrices
de tierra en un Arguidol bajo cultivo de soya (Colonia Ensayo, Entre Rios). Revista Ciencia Agropecuaria. 19(1-2):39-50.
Matienzo, BY; Alfonso-Simonetti, J; Vásquez, ML; De la Masa, AR; Matamoros, TCh; Díaz, FY. 2015. Diversidad de grupos funcionales
de la fauna edáfica y su relación con el diseño y manejo de tres sistemas de cultivos. Fitosanidad. 19(1):45-55.
Mendieta-Araica, Bryan. 2011. Moringa oleifera as an alternative fodder for dairy cows in Nicaragua. Doctoral Thesis. Swedish University
of Agricultural Sciences. Uppsala. p 17-25.
Moreno, C. 2001. Métodos para medir la biodiversidad. M & T-Manuales y Tesis SEA, Vol. 1. Zaragoza, 84 p.
Nieto, GL; Valencia, TF; Giraldo, DR. 2013. Bases pluriepistemológicas de los estudios en agroecología. Entramado. 9(1):204-211.
Padoa, L; Eberthardt, DN; Marchanāo, RL; Corbeels, M; Scopel, E; Muraoka, T. 2012. Soil macrofauna and its relationship with
Carbon and Nitrogen contents under conservation agriculture systems in the Cerrado of Unai. FERTBIO. 5 p.
Palada, M; Chang, Lien-Chung. 2003. Suggested cultural practices for moringa. Asian Vegetable Research and Development Center.
6 p.
Rendón, PS; Artunduaga, LF; Ramirez, PR; Quiroz, GJ; Leyva, RE. 2011. Los macroinvertebrados como indicadores de la calidad del
suelo en cultivos de Mora, Pasto y Aguacate. Revista Facultad Nacional de Agronomía. 64(1)10.
Roldan, G. 1988. Guía para el estudio de macroinvertebrados acuáticos del Departamento de Antioquia. Centro de Investigaciones.
Universidad de Antioquia. Bogotá, Colombia. 215 p.
Sermeño, Ch.J.M. 2013. Bío ecología e identificación de las familias de termitas (Blattaria: Isoptera) presentes en El Salvador. Revista
Bioma. 22(1):9-14.
Socorrás, A; Izquierdo, I. 2014. Evaluation of agroecological systems through biological indicators of the soil quality: edaphic mesofauna.
Pastos y forrajes. 37(1):109-114.
Turner, M; Garner, R. (Ed). 1991. Quantitative methods in landscapes ecology. Ecological Studies 82. Springer-Verlag, New York.
536 p.
Vega, MA; Herrera, RS; Rodríguez, GA; Sánchez, S; Lamela, L; Santana, AA. 2014. Evaluación de la macrofauna edáfica en un sistema
silvopastoril en el Valle del Cauto, Cuba. Revista Cuba de Ciencia Agrícola. 48(2):189-193.
Vieira Da Cunha, NF; Fernandes, C.M.E; Almeida, P.G.H; Pereira, MG; Dos Santos Leles, PS. 2012. Soil fauna as an indicator of soil
quality in forest stands, pasture and secondary forest. Revista Brasileira de Ciencia do Solo. 36(5):1407-1417.
Zerbino, MS. 2010. Evaluación de la macrofauna del suelo en rotaciones cultivos-pasturas con laboreo convencional. Acta Zoológica
Mexicana. N°2:189-202.
|
Recibido: 29 de mayo 2017
Aceptado: 7 de agosto del 2017
|
 |