Recursos Naturales y Ambiente
Comunidades arbóreas asociadas a la elevación en la microcuenca El Zapote, Matagalpa, Nicaragua
Tree assemblages associated to elevation at El Zapote, watershed, Matagalpa, Nicaragua
La Calera
Universidad Nacional Agraria, Nicaragua
ISSN: 1998-7846
ISSN-e: 1998-8850
Periodicidad: Semestral
vol. 21, núm. 36, 2021
Recepción: 06 Julio 2020
Aprobación: 01 Febrero 2021

Resumen: Entender la relación entre la flora arborescente con su ambiente es crucial como base para cualquier programa de restauración y conservación. Este trabajo plantea determinar las comunidades arbóreas asociadas a un gradiente de elevación, seleccionar las especies indicadoras de cada comunidad y relacionar la composición de especie con el estado de conservación del bosque. Para lograr los objetivos se establecieron 32 parcelas rectangulares de 20 m x 50 m de forma aleatoria en el área de estudio, el cual está representado por un bosque transicional relativamente conservado en la microcuenca El Zapote, Matagalpa. Dentro de la parcela se registró la abundancia y tipo de especie de todos los árboles ≥10 cm de diámetro normal (1.30 m del suelo) y se registró la elevación (m) en la que se ubicaba cada parcela. Mediante estadística multivariada se dilucidaron dos comunidades arbóreas asociadas a un gradiente de elevación; las especies Guazuma ulmifolia y Cordia alliodora son indicadoras de la comunidad ubicada a elevación de 717.2±30.0 m y las especies Daphnopsis americana, Stemmadenia donnell-smithii, Guettarda macrosperma, Trichilia trifolia, Pouteria sapota, Brosimum alicastrum, Brunellia mexicana, Inga vera, Stemmadenia pubescens y Lonchocarpus phlebophyllus son indicadoras de la comunidad ubicada a elevación de 856.3±41.1 metros. Es probable que en la parte alta de la microcuenca aún se conserven remanentes del bosque autóctono del área de estudio. Palabra claves: composición, elevación, flora arborescente.
Palabras clave: composición, elevación, flora arborescente.
Abstract: Understanding the relationship between tree flora and its environment is important as base for any restoration and conservation program. This work aims to determine the tree communities associated with an elevation gradient; select indicator species from each community; and relate the species composition to the forest's conservation status. To achieve the objectives, 32 rectangular plots (20 m x 50 m) were randomly established at the study area, which is represented by a relatively preserved transitional forest in El Zapote micro-watershed, Matagalpa. Within the plot, the abundance and species type of all trees ≥10 cm normal diameter (1.30 m from the ground) were recorded; the elevation (m) for each plot was also obtained. Using multivariate statistics, two tree communities associated with an elevation gradient were elucidated; the Guazuma ulmifolia and Cordia alliodora species are indicators of the community located at an elevation of 717.2±30.0 m; and the species Daphnopsis americana, Stemmadenia donnell-smithii, Guettarda macrosperma, Trichilia trifolia, Pouteria sapota, Brosimum alicastrum, Brunellia mexicana, Inga vera, Stemmadenia pubescens and Lonchocarpus phlebophyllus are indicative of the community located at elevation of 856.3±41. It is probable that native forest remnants are still preserved in the upper part of the micro-watershed.
Keywords: Composition, elevation, tree flora.
Definir las comunidades arbóreas en un bosque es conocimiento básico para cualquier iniciativa de conservación de este, en especial si incluye el componente de restauración o rehabilitación (Vargas, 2007). Las comunidades arbóreas tienen una composición de especie que está directamente relacionada con variables ambientales que a escala de paisaje están influenciadas por diferentes gradientes y combinaciones de gradientes (Chai et al., 2016; Krishnadas et al., 2016). La elevación es una variable de importancia en la definición de la composición de estas comunidades (Toledo-Garibaldi y Williams-Linera, 2014; Cuyckens et al., 2015; Jiménez et al., 2016).
A lo largo del gradiente de elevación se definen y combinan otros gradientes (de variables asociadas al suelo y al clima) que en conjunto conforman las condiciones edafoclimáticas en donde se organiza la vegetación arbórea. Es muy conocida la influencia de la elevación en los factores suelo y precipitación (Johansson y Chen, 2003; Sánchez et al., 2005; Sanchez-Moreno et al., 2014) y por ende en la biodiversidad de plantas (Kessler et al., 2012; Irl et al., 2015; Qanbari y Akbar, 2015). Los programas de restauración o rehabilitación de ecosistemas forestales contemplan el conocer esta complejidad de interacciones para alcanzar la meta de recuperar las comunidades arbóreas autóctonas organizadas a lo largo de gradientes ambientales (Vargas, 2007).
Para aportar al conocimiento de las comunidades arbóreas en el área de estudio y su relación con la elevación (gradientes ambientales asociados a esta) y sentar las bases de futuras investigaciones en pro de la conservación del bosque de la microcuenca El Zapote, son objetivos de este estudio el determinar las comunidades arbóreas asociadas a un gradiente de elevación, seleccionar las especies indicadoras de cada comunidad y relacionar la composición de especie con el estado de conservación del bosque.
El aporte de este trabajo es, además de definir las comunidades arbóreas en el gradiente de elevación, determinar especies indicadoras de cada comunidad, las cuales sean útiles para el biomonitoreo del estado de conservación del bosque definido por las comunidades. Los cambios en la abundancia y frecuencia de ocurrencia de estas especies podrían, en la escala de tiempo, estar asociado con perturbaciones (incluyendo cambio climático global o local, influencia humana y eventos naturales) particulares en la composición de especies de la comunidad vegetal que representan (especies indicadoras) y por lo tanto del bosque completo.
MATERIALES Y MÉTODOS
Descripción del sitio
La microcuenca El Zapote se ubica entre los municipios Terrabona y San Dionisio del departamento de Matagalpa en las coordenadas 12°42'49" N, 85°53'46" O y 12°46'45” N, 85°56'13" O. Posee un área total de 1 772 hectáreas de los cuales aproximadamente un 28 % están cubiertos por bosque. La vegetación típica es de zona transicional entre bosque seco tropical y bosque húmedo con elevaciones entre los 700 y 1 200 metros (Meyrat, 2006). La temperatura oscila entre 21 y 24 °C y precipitaciones entre 1 200 y 1 800 mm anuales. En la microcuenca predominan suelos de los órdenes Molisoles y Vertisoles. La principal actividad productiva en el área es el cultivo de café con sombra en sistemas agroforestales.
Toma de datos
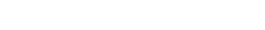
Se establecieron un total de 32 parcelas con dimensión de 20 x 50 m (0.1 ha), las que se ubicaron de forma aleatoria con el uso de la herramienta Random en ArcGIS 10.2, seleccionando el uso de suelo “bosque” como área potencial para la ubicación, el que corresponde un área de 500 ha. De tal forma que el esfuerzo de muestreo fue de 0.64 %. Las imágenes satelitales utilizadas para definir el área boscosa fue la del sensor Landsat 8 e imágenes de Google Earth. En la Figura 1 se muestra un mapa del área de estudio, las áreas oscuras representan al bosque en donde se establecieron aleatoriamente las 32 unidades de muestreo, representadas por los puntos.
Dentro de las parcelas se registraron todos los individuos de especies arbóreas ≥10 cm de diámetro normal (1.30 m del suelo). Las principales variables tomadas fueron el tipo de especie, el número de individuos y la elevación en metros. Adicionalmente se registró el diámetro (cm) y la altura (m) de cada árbol con propósito descriptivo general. Las identificaciones se realizaron en campo utilizando las guías ilustradas de Salas (1993), Cordero y Boshier (2003), Quezada et al. (2010), Meyrat et al. (2012). Las especies no identificadas en campo fueron colectadas e identificadas en el Herbario Juan de Dios Rostrán de la Universidad Nacional Agraria y el Herbario Nacional de la Universidad Centroamericana. La taxonomía que se siguió fue la propuesta por Stevens et al., (2001).
Análisis de la información
Se realizó una descripción general de la flora arborescente que consistió en hacer recuento de abundancia total, conteo del número de especies, distribución taxonómica, selección de especies y familias más representativas, descripción general de estructura del bosque en términos de clases diamétricas y de altura de los árboles.
Seguidamente se definieron las comunidades en base a la composición de las especies determinadas en el área de estudio. En estos análisis solamente se incluyeron 35 especies (36.5 % del total), las que tenían abundancias mayores o iguales a 10 individuos y al menos ocurrieron en más de tres parcelas, a fin de poder controlar datos atípicos, valores influencivos y seleccionar las especies que aportaban mayor explicación al modelo.
Para lo anterior se utilizaron análisis estadísticos multivariados, entre ellos el Análisis de Conglomerado con distancia Euclideana y con el método de Conglomerado de Varianza Mínima de Ward, utilizando una matriz de datos de 32 filas y 35 columnas, correspondiendo a las parcelas y a las especies con sus valores de abundancia. Con este análisis se definieron los grupos (también llamados comunidades) de parcelas asociados en base a la composición de las especies. La relación entre las comunidades definidas y la variable Elevación se exploraron en una ordenación llamada Escalamiento Multidimensional No-Métrico, (McCune y Grace, 2002; Borcard et al., 2011) o NMS por sus siglas en inglés, con distancia Bray-Curtis. El análisis de conglomerado y la ordenación NMS se realizaron con el programa R 3.2.2. (R Core Team, 2015) y los paquetes MASS (Ripley et al., 2018) y Vegan (Oksanen et al., 2018).
Para explorar diferencias significativas en términos de composición entre los grupos resultantes se utilizó el Procedimiento de Permutación Multirespuesta (MRPP por sus siglas en inglés) (McCune y Grace, 2002) con distancia Euclideana (Pythagorean). El MRPP provee un valor A útil para determinar cuan similar son las unidades de muestreo entre cada grupo. Cuando la heterogeneidad entre grupos fue igual a lo esperado por chance A = 0; si las observaciones dentro de los grupos fueran idénticas, A = 1; cuando hay más heterogeneidad entre grupos que la esperada por chance, A < 0. Adicionalmente, un valor de p (probabilidad) es proveído para evaluar la probabilidad de observar diferencias por chance.
Las especies ecológicamente importantes fueron determinadas con el Análisis de Especies Indicadoras (ISA) (McCune y Grace, 2002). Este análisis combina matemáticamente la abundancia relativa y la frecuencia relativa de las especies dentro de un grupo dado y la fiabilidad de ocurrencia de esas especies en tales grupos. El análisis proporciona un valor de indicación (VI) y un valor de p para cada especie utilizando el procedimiento de permutación de Monte Carlo con 4 999 aleatorizaciones. El nivel de significancia utilizado en este trabajo fue de 0.05. Los análisis MRPP e ISA se realizaron en el programa PCORD 6.08 (McCune y Mefford, 2011).
RESULTADOS Y DISCUSIÓN
Descripciones generales de la flora arborescente
En el área muestreada se contaron un total de 1 186 individuos, distribuidos en 96 especies, 74 géneros y 35 familias. Las especies más abundantes según el número de individuos fueron en principio: Trichilia trifolia (115 individuos) y Guazuma ulmifolia (111), seguidos de Lonchocarpus chiangii (75), Cordia alliodora (57), Inga vera (56), Daphnopsis americana (46), Guettarda macrosperma (40), Thouinidium decandrum (38) y Platymiscium parviflorum (34). El resto de las especies fueron registradas con menos de 30 individuos. La familia más abundante según el número de especies fue Fabaceae (23 especies) seguida de Meliaceae (6) y Rubiaceae (5). El resto de las familias estaban representadas por menos de cuatro especies.
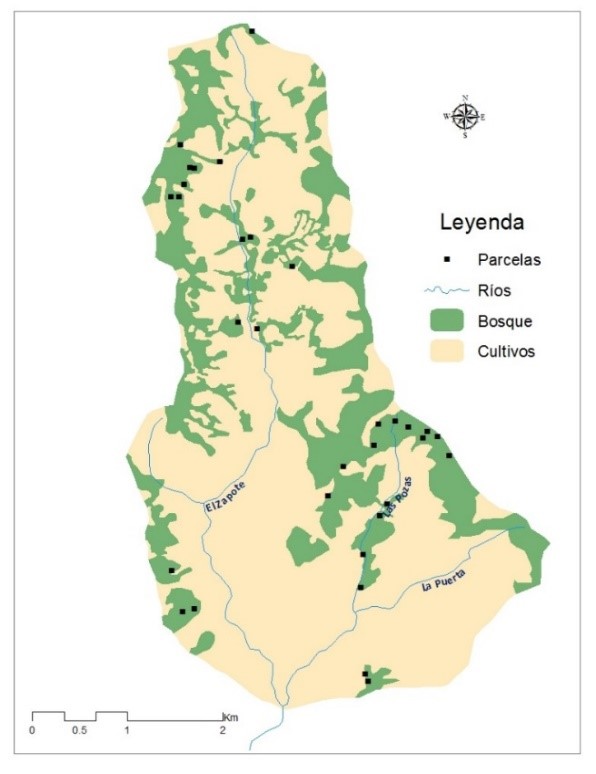
De manera general el diámetro promedio ± error estándar de los árboles muestreados fue de 21.7 ± 0.5 cm. A como se muestra en la Figura 2, la mayoría (84 %) de los individuos de flora arborescente muestreados pertenecían a los rangos de diámetro 10 – 20 cm seguidos de 20 – 30 cm; siendo representativas (según el número de individuos) en estos rangos de diámetro las especies más abundantes antes mencionadas. Una minoría (0.5 %) se encuentra entre los rangos 80 – 130 cm, representado por las especies Guettarda macrosperma, Terminalia oblonga, Ficus insipida, Brosimum alicastrum, Albizia guachapele y Enterolobium cyclocarpum.
En cuanto a la altura, los árboles tenían altura promedio ± error estándar de 9.4 ± 0.1 m. La mayoría (70 %) de los individuos de flora arborescente muestreados se encontraban en los rangos de altura de 7 – 12 m. Una minoría (0.8 %) se encuentra entre los rangos 20 – 25 m, entre las especies que se encuentran en este rango están: Cecropia peltata, Cedrela odorata, Lonchocarpus chiangii y Terminalia oblonga.
La distribución de las dos variables es asimétrica y la mayor densidad de ocurrencia se concentra en los menores valores, este tipo de distribución es evidencia de un bosque en desarrollo con mayor abundancia de individuos de porte pequeño y muy raros individuos de porte grande (González-Rivas, 2005; Garmendia et al., 2008; Zamora, 2010; Obando y Oliva, 2011; Blanco, 2017; Bustillo, 2017).
Definición de comunidades basado en composición
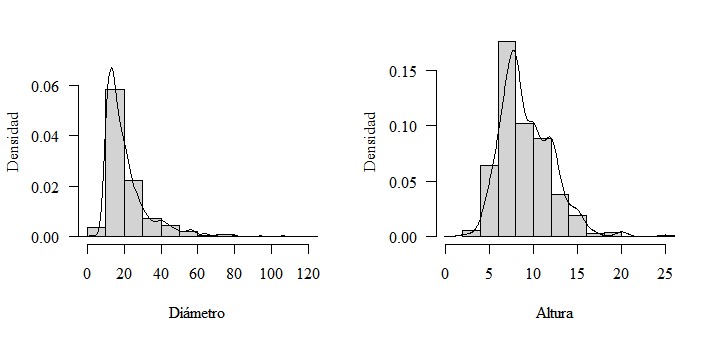
Como análisis exploratorio preliminar para la definición de comunidades basado en la composición se utilizó el Análisis de Conglomerado, con el cual se definieron claramente dos grupos de aglomeración de las parcelas de muestreo, el primer grupo agrupando 19 parcelas y el segundo 13 parcelas. En la Figura 3 se aprecian en análisis con los dos grupos, el grupo 1 a la izquierda y el grupo 2 a la derecha. Con la formación de los dos grupos heterogéneos se sospecha que la vegetación está constituida por dos comunidades arbóreas, el siguiente paso fue confirmar la existencia de esas comunidades y su asociación con la variable elevación.
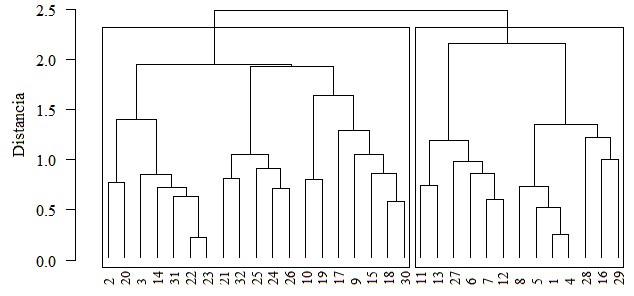
La interacción entre las dos comunidades (grupos) formadas mediante el Análisis de Conglomerado y la variable Elevación (m) fueron visualizadas por medio de un Escalamiento Multidimensional No-Métrico (NMS) con distancia Bray-Curtis. Para esta ordenación se obtuvo una solución bidimensional con 0.25 de estrés y 20 iteraciones.
En la ordenación (Figura 4) se muestran tres componentes: 1) Las parcelas representadas con puntos huecos; 2) Las especies con las etiquetas de sus nombres codificados y 3) El vector correspondiente a los valores de la variable Elevación. Los polígonos encierran a los sitios agrupados según el Análisis de Conglomerado anterior (Figura 3). La ordenación NMS evidencia que existen dos comunidades separadas formando parte de la composición florística de toda el área de estudio y también que esta divergencia de comunidades podría estar asociada a la variable Elevación.
Las diferencias en las comunidades son notorias en el NMS pues los polígonos solo se superponen en una pequeña región de estos, haciendo notoria la sospecha de que en realidad son agrupaciones vegetales diferentes (en términos de composición). También es notorio que el eje X del gráfico podría está representado por el gradiente de Elevación (determinado por el vector) el que aumenta sus valores hacia la izquierda, por consiguiente, el polígono de la izquierda (el grupo 2 en la Figura 3) contiene las parcelas, y por ende las especies, más asociadas a valores altos de la elevación; y en contraposición el polígono de la derecha (el grupo 1 en la Figura 3) contiene las parcelas, y las especies, más asociadas a valores bajos de la elevación.
En el NMS se pueden visualizar algunas especies que están asociadas a valores altos de elevación, como DAPHAMER, STEMDONN, BRUNMEXI, STEMPUSE (nombres completos en Cuadro 1) entre otras, y algunas especies que están asociadas a valores bajos de elevación como GUAZUMI, CEDREODR, CORDIALLI, PLATPARV entre otras; sin embargo, el NMS es un análisis exploratorio global para visualizar las relaciones, en términos de inferir diferencia entre las dos comunidades y de evaluar la verdadera asociación de las especies en cada comunidad se utilizó el Procedimiento de Permutación Multirespuesta (MRPP) y el Análisis de Especies Indicadoras (ISA).
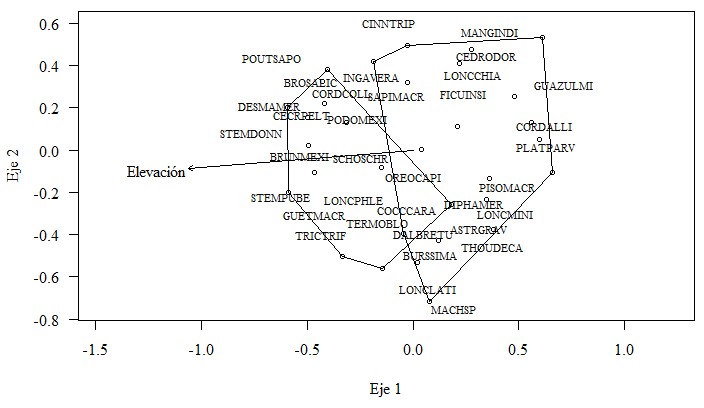
Utilizando en MRPP, se comparó la composición de especies de las dos comunidades (grupos) formados y visualizados en el NMS. Como resultado se confirma que existe diferencias altamente significativas en términos de composición (A=0.053, p=0.000025). En base a este resultado se puede concluir que realmente el bosque en el sitio de estudio en general está definido por dos comunidades y que esas diferencias pueden estar influenciadas con la elevación, ya que el valor promedio de la elevación en las parcelas de la comunidad asociada positivamente a dicha variable es de 856.3±41.1 m (media ± error estándar) y el valor promedio de la elevación en las parcelas de la comunidad asociada negativamente a dicha variable es de 717.2±30.0 m. La diferencia de la elevación comparada entre los grupos de parcelas es significativa (p=0.01).
Selección de especies indicadoras por comunidad (grupo)
Con el MRPP se confirmó la diferencia entre las dos comunidades en términos de composición, esto también fue notorio visualmente en el NMS; seguidamente mediante el Análisis de Especies Indicadoras se determinaron las especies en particular que son representativas (en cuanto a abundancia y frecuencia relativa) de cada comunidad y que por lo tanto, son ecológicamente importantes.
El Cuadro 1 muestra el resultado del Análisis de Especies Indicadoras para cada una de las comunidades definidas en la Figura 3 y visualizadas en la Figura 4. El resultado general de la prueba de Monte Carlo resultó ser significativo (p=0.0002). Las especies Guazuma ulmifolia (76.7) y Cordia alliodora (65.0) fueron las que mayor Valor de Indicación (VI) obtuvieron para el grupo 1 (visualizado como el polígono de la derecha en la Figura 4), con valores de p altamente significativo y muy significativo respectivamente (p=0.0008 y p=0.0030). Siendo estas las especies ecológicamente importantes en esta comunidad que ocurre en elevación promedio de 717.2±30.0 m. El resto de las especie de ese grupo (14) resultaron con Valor de Indicación menores a 35 y con p≥0.05.
| Nombre científico | Código | Grupo | VI | p | |
| Guazuma ulmifolia | GUAZULMI | 1 | 76.7 | 0.0008 | *** |
| Cordia alliodora | CORDALLI | 1 | 65.0 | 0.0030 | ** |
| Thouinidiumdecandrum | THOUDECA | 1 | 34.5 | 0.0670 | |
| Diphysa americana | DIPHAMER | 1 | 23.8 | 0.1946 | |
| Lonchocarpus chiangii | LONCCHIA | 1 | 44.3 | 0.2084 | |
| Astronium graveolens | ASTRGRAV | 1 | 23.5 | 0.2244 | |
| Coccoloba caracasana | COCCCARA | 1 | 15.8 | 0.2531 | |
| Cedrela odorata | CEDRODOR | 1 | 23.7 | 0.2537 | |
| Machaerium Sp. | MACHSP | 1 | 16.3 | 0.3479 | |
| Platymiscium parviflorum | PLATPARV | 1 | 22.9 | 0.4491 | |
| Lonchocarpus minimiflorus | LONCMINI | 1 | 16.2 | 0.4617 | |
| Mangifera indica | MANGINDI | 1 | 10.5 | 0.5047 | |
| Lonchocarpus latifolius | LONCLATI | 1 | 11.0 | 0.6927 | |
| Pisonia macranthocarpa | PISOMACR | 1 | 12.2 | 0.8174 | |
| Bursera simaruba | BURSSIMA | 1 | 12.8 | 0.9022 | |
| Ficus insípida | FICUINSI | 1 | 8.70 | 0.9302 | |
| Daphnopsis americana | DAPHAMER | 2 | 73.4 | 0.0002 | *** |
| Stemmadenia donnell-smithii | STEMDONN | 2 | 53.8 | 0.0008 | *** |
| Guettarda macrosperma | GUETMACR | 2 | 59.1 | 0.0048 | ** |
| Trichilia trifolia | TRICTRIF | 2 | 52.2 | 0.0062 | ** |
| Pouteria sapota | POUTSAPO | 2 | 44.7 | 0.0076 | ** |
| Brosimum alicastrum | BROSALIC | 2 | 36.0 | 0.0176 | * |
| Brunellia mexicana | BRUNMEXI | 2 | 35.7 | 0.0200 | * |
| Inga vera | INGAVERA | 2 | 47.2 | 0.0236 | * |
| Stemmadenia pubescens | STEMPUBE | 2 | 30.8 | 0.0236 | * |
| Lonchocarpus phlebophyllus | LONCPHLE | 2 | 40.4 | 0.0314 | * |
| Cecropia peltata | CECRPELT | 2 | 29.7 | 0.0786 | |
| Cordia collococca | CORDCOLL | 2 | 20.3 | 0.3517 | |
| Podopterus mexicanus | PODOMEXI | 2 | 29.8 | 0.4269 | |
| Sapium macrocarpum | SAPIMACR | 2 | 21.8 | 0.4567 | |
| Terminalia oblonga | TERMOBLO | 2 | 12.2 | 0.5085 | |
| Oreopanax capitatus | OREOCAPI | 2 | 17.3 | 0.6025 | |
| Schoepfia schreberi | SCHOSCHR | 2 | 13.7 | 0.7407 | |
| Cinnamomum triplinerve | CINNTRIP | 2 | 14.9 | 0.7720 | |
| Dalbergia retusa | DALBRETU | 2 | 13.0 | 0.7844 | |
Las especie Daphnopsis americana (73.4) y Stemmadenia donnell-smithii (53.8) fueron las que mayor VI obtuvieron del grupo 2 (visualizado como el polígono de la izquierda en la Figura 4), con valores de p altamente significativos respectivamente (p=0.0002 y p=0.0008). A D. americana y S. donnell-smithii le siguen tres especies también con VI altos y p muy significativos (p=0.0048, p=0.0062 y p=0.0076): Guettarda macrosperma (59.1), Trichilia trifolia (52.2) y Pouteria sapota (44.7) y cinco especies con VI altos y medios, y p significativos (p=0.0176, p=0.0200, p=0.0236, p=0.0236 y p=0.0314): Brosimum alicastrum (36.0), Brunellia mexicana (35.7), Inga vera (47), Stemmadenia pubescens (30.8) y Lonchocarpus phlebophyllus (40.4). Siendo estas las especies ecológicamente importantes en esta comunidad que ocurre en elevación promedio de 856.3±41.1 m. El resto de las especie de ese grupo (9) resultaron con VI menores a 30 y con p≥0.05.
La comunidad asociada negativamente con la elevación (grupo 1) ocurren principalmente en bosques con estados sucesionales tempranos e influenciados por las actividades humanas, en especial G. ulmifolia, una especie pionera de bosques jóvenes, asociada con bosques perturbados y pastizales (Francis, 1991; Stevens et al., 2001) de la zona seca o transicional. La otra especie con alto VI en este grupo fue C. alliodora, una especie muy utilizada por los productores por su alto valor como madera y su potencial para ser combinado con cultivos (sistemas agroforestales) en los que se aprecia su copa abierta y rala, y su capacidad de autopoda (Cordero y Boshier, 2003). Otras especies que evidencian fuerte influencia humana en el área dominada por la comunidad representada por G. ulmifolia están: Coccoloba caracasana y Mangifera indica. De hecho, en la parte media y baja de la microcuenca es usual encontrar sistemas agroforestales de café con sombra (Garmendia y Herrera, 2018), de tal forma que probablemente los bosques aledaños a estos sistemas hayan sufrido cierta influencia humana.
Las especies con alto VI que forman parte de la comunidad asociada positivamente con la elevación (grupo 2) ocurren principalmente en ecosistemas naturales de la zona y asociadas a bosques con relativa poca perturbación. Daphnopsis americana es una especie bien distribuida en bosques siempreverdes en el norte del país; S. donnell-smithii es una especie típica de bosque seco con cierta sombra; G. macrosperma y T. trifolia ocurren principalmente en bosques estacionales, de galería o deciduos y a veces siempreverdes; P. sapota aunque es popularmente cultivado, es frecuente en bosques húmedos; B. alicastrum es común en bosques deciduos y siempreverdes, así como en riberas en zonas semiárida; B. mexicana es una especie rara de nebliselva de la zona central; I. vera una especie muy utilizada en sistemas agroforestales, pero que también ocurre en ecosistemas naturales secos y húmedos; S. pubescens común en bosques secos y caducifolios y L. phlebophyllus común en bosques caducifolios en la zona pacífica y norcentral (Stevens et al., 2001; Cordero y Boshier, 2003).
CONCLUSIONES
La composición del bosque transicional de la microcuenca El Zapote está representado por dos comunidades arbóreas asociadas a un gradiente de elevación.
Las especies indicadoras de las zonas más bajas fueron, en orden de importancia: Guazuma ulmifolia y Cordia alliodora; en cambio, en las zonas más altas fueron, en orden de importancia: Daphnopsis americana, Stemmadenia donnell-smithii,Guettarda macrosperma, Trichilia trifolia, Pouteria sapota, Brosimum alicastrum, Brunellia mexicana, Inga vera, Stemmadenia pubescens y Lonchocarpus phlebophyllus.
Debido a la presencia de especies arbóreas que son ecológicamente importantes en la parte alta de la microcuenca, se puede deducir que en esta zona se conservan remanentes de bosques naturales con poco grado de intervención; opuesto a lo que ocurre en la parte baja, donde las especies son representativas de bosques secundarios y alterados, incluyendo especies introducidas por el ser humano.
Referencias
Blanco, B. (2017). Dinámica Poblacional en una Cronosecuencia de Bosque Seco Tropical, Parque Nacional Santa Rosa, Guanacaste, Costa Rica [Tesis de grado, Tecnológico de Costa Rica]. Repositorio Tecnológico de Costa Rica. http://hdl.handle.net/2238/9241
Borcard, D., Gillet, F. y Legendre, P. (2011). Numerical Ecology with R. Springer
Bustillo, I. (2017). Composición, Estructura y Dinámica de las Especies Arbóreas del Bosque de Galería de la Reserva Hídrica Forestal, Río Malacatoya, 2015-2016 [Tesis de grado, Universidad Nacional Agraria]. Repositorio Institucional Universidad Nacional Agraria. https://repositorio.una.edu.ni/3577/1/tnk10b982c.pdf
Chai, Z., Fan, D. y Wang, D. (2016). Environmental factors and underlying mecanisms of tree community assamblages on pine-oak mixed forests in the Qinling Mountains, China. Journal of Plant Biology, 59, 347-357. https://doi.org/10.1007/s12374-015-0503-0
Cordero, J. y Boshier, D. H. (ed.). (2003). Árboles de Centroamérica, un Manual para Extensionistas. Centro Agronómico Tropical de Investigación y Enseñanza.
Cuyckens G, A. E., Malizia, L. R. y Blundo, C. (2015). Composición, diversidad y estructura de comunidades de árboles en un gradiente altitudinal de selvas subtropicales de montaña (Serranías de Zapla, Jujuy, Argentina). Madera y Bosques, 21(3), 137-148. https://doi.org/10.21829/myb.2015.213463
Francis, J. K. (1991). Guazuma ulmifolia Lam. Guácima. SO-ITF-SM-47. U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station.
Garmendia, G. y Herrera J. (2018). Relación entre la composición de la flora arbórea y variables ambientales, dasométricas y silviculturales en dos ecosistemas de la microcuenca “El Zapote”, Terrabona - San Dionisio, Matagalpa, Nicaragua [Tesis de grado, Universidad Nacional Agraria]. Repositorio Institucional Universidad Nacional Agraria. https://repositorio.una.edu.ni/3734/1/tnk10g233.pdf
Garmendia, M., Quezada, J. B., Armas, H. y Meneces, E. (2008). Composición, diversidad, estructura e importancia de las especies arbóreas y palmas del bosque seco de la finca “Rosita”, Reserva Natural Estero Padre Ramos, Chinandega, Nicaragua. La Calera, 8(11), 66-73. https://lacalera.una.edu.ni/index.php/CALERA/article/view/125/125
González-Rivas, B. (2005). Tree species diversity and regeneration of tropical dry forest in Nicaragua [Tesis doctoral, Swedish University of Agricultural Sciences]. Open access publications in the SLU publication database. https://pub.epsilon.slu.se/837/1/FinalFinalSummary.pdf
Irl, S., Harter, D., Steinbauer, M., Gallego, D., Fernández-Palacios, J., Jentsch, A. y Beierkuhnlein, C. (2015). Climate vs. topography - spatial patterns of plant species diversity and endemism on a high-elevation island. Journal of Ecology, 103, 1621–1633. https://doi.org/10.1111/1365-2745.12463
Jiménez, L., Capa-Mora, D., Quichimbo, P., Mezquida, E., Benito, M. y Rubio, A. (2016). Influencia de las características ambientales en la composición florística de un bosque siempreverde piemontano. Bosque Latitud Cero, 6(2), 88-103. https://revistas.unl.edu.ec/index.php/bosques/article/view/232/215
Johansson, B. y Chen, D. (2003). The influence of wind and topography on precipitation distribution in Sweden: Statistical analysisi and modeling. International Journal of Climatology, 23(12), 1523-1535
Kessler, M., Grytnes, J., Halloy, S., Kluge, J., Krömer, T., León, B., Marcía, M. y Young, K. (2012). Gradientes de Diversidad Vegetal: Patrones y Procesos Locales. En S.K. Herzog, R. Martínez, P.M. Jørgensen, H. Tiessen. (Eds.) Cambio Climático y la Biodiversidad en los Andes Tropicales (pp. 235-253). Inter-American Institute of Global Change Research (IAI), Scientific Committee on Problems of the Environment (SCOPE). https://jcmendo.com/wp-content/uploads/2018/docs-publi-cursos/documentacion-tecnica/Cambio%20Climatico%20y%20Biodiversidad%20en%20los%20Andes%20Tropicales%20Foundation%20Mc%20Arthur.pdf
Krishnadas, M., Kumar, A. y Comita, L.S. (2016). Environmental gradients structure tropical tree assamblages at the regional scale. Journal of Vegetation Science, 27(6), 1117-1128. https://doi.org/10.1111/jvs.12438
McCune, B. y Grace, J. B. (2002). Analysis of Ecological Communities. MjM Software Design
McCune, B. y M. J. Mefford. (2011). PC-ORD. Multivariate Analysis of Ecological Data. (Version 6.08) [Software].
Meyrat, A. (2006). Descripción de Ecosistemas y Formaciones Vegetales de Nicaragua. Ministerio del Ambiente y los Recursos Naturales.
Meyrat, A., Quezada, J. B. y Garmendia, M. (2012). Árboles y Arbustos Predominantes de Nicaragua. Franklin Ruíz M. NOC Graphic & Web Design.
Obando, R. y Oliva, E. (2011). Estudio de la dinámica poblacional de diez especies arbóreas en un período 2006-2007 en el bosque seco secundario en la Comarca La Chipopa, Nandaime, Granada [Tesis de grado, Universidad Nacional Agraria]. Repositorio Institucional Universidad Nacional Agraria. https://repositorio.una.edu.ni/1157/1/tnk10o12.pdf
Oksanen, J., Guillaume, F., Friendly, M., Kindt, R., Legendre, P., McGlinn, D. y Wagner, H. (2018). Community Ecology Package “vegan”. (Version 2.5-2) [Software]. https://cran.r-project.org/
Qanbari, V. y Akbar, A. (2015). The relationship between elevation, soil properties and vegetation cover in the Shorb-Ol-Ain watershed of Yazd. Journal of Biodiversity and Environmental Sciences, 6(5), 49-56. https://www.innspub.net/wp-content/uploads/2015/05/JBES-Vol6No5-p49-56.pdf
Quezada, J. B., Garmendia, M. y Meyrat, A. (2010). Especies de árboles del Arboretum Alain Meyrat (Vol. 1). Universidad Nacional Agraria. https://repositorio.una.edu.ni/2462/1/nf70q5v1.pdf
R Core Team. (2015). R: A language and environment for statistical computing. (Version 3.2.2) [Software]. http://www.R-project.org/
Ripley B., Venables, B., Bates, D., Hornik, K., Gebhardt, A. y Firth, D. (2018). Support Functions and Datasets for Venables and Ripley's MASS. (Version 7.3-50) [Software]. https://cran.r-project.org/
Salas, J. B. (1993). Arboles de Nicaragua. IRENA.
Sánchez, B., Ruiz, M. y Ríos, M. (2005). Materia orgánica y actividad biológica del suelo en relación con la altitud, en la cuenca del río Maracay, estado Aragua. Agronomía Tropical, 56(4), 507-534. http://ve.scielo.org/pdf/at/v55n4/art04.pdf
Sanchez-Moreno, J. F., Mannaerts, C. M. y Jetten, V. (2014). Influence of topography on rainfall variability in Santiago Island, Cape Verde. International Journal of Climatology, 34, 1081-1097. https://doi.org/10.1002/joc.3747
Stevens, W., Ulloa, C., Pool, A. y Montiel, O. (ed.). (2001). Flora de Nicaragua (Tomo I, II y III). Missouri Botanical Garden.
Toledo-Garibaldi, M. y Williams-Linera, G. (2014). Tree diversity patters in successive vegetation type along an elevation gradient in the Mountains of Eastern Mexico. Ecological Research, 29(6), 1097-1104. https://doi.org/10.1007/s11284-014-1196-4
Vargas, O. (ed.). (2007). Guía Metodológica para la restauración ecológica del bosque altoandino. Universidad Nacional de Colombia. http://observatorio.epacartagena.gov.co/wp-content/uploads/2016/06/guia-metodologica-restauracion-ecologica.pdf
Zamora, M. (2010). Caracterización de la flora y estructura de un bosque transicional húmedo a seco, Miramar, Puntarenas, Costa Rica [Tesis de grado, Tecnológico de Costa Rica]. Repositorio Tecnológico de Costa Rica. http://hdl.handle.net/2238/3017
Notas de autor