Ciencia de las Plantas
Capacidad inhibitoria de productos biológicos y sintéticos sobre el crecimiento micelial del oomiceto Phytophthora infestans causante del tizón tardío
Inhibitory capacity of biological and synthetic products on the mycelial growth of the oomycete Phytophthora infestans, the causal agent of late blight


Capacidad inhibitoria de productos biológicos y sintéticos sobre el crecimiento micelial del oomiceto Phytophthora infestans causante del tizón tardío
La Calera, vol. 26, núm. 46, pp. 24-34, 2026
Universidad Nacional Agraria

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Recepción: 13 Enero 2026
Aprobación: 29 Abril 2026
Resumen: El tizón tardío, causado por el oomiceto Phytophthora infestans, constituye uno de los principales factores limitantes en la producción de papa (Solanum tuberosum L.) en Nicaragua. El objetivo de esta investigación fue evaluar la capacidad inhibitoria in vitro de productos biológicos y sintéticos para el manejo de Phytophthora infestans (Tizón tardío). Se utilizaron aislados del patógeno provenientes de cuatro fincas de las comunidades El Robledal y El Zacatón, ubicadas en el departamento de Estelí, al Norte de Nicaragua. El aislamiento de Phytophthora infestans a partir de suelo, se realizó mediante el método de cultivo trampa, utilizando fragmentos de hojas de clavel (Dianthus caryophyllus) como material vegetal trampa, posteriormente fueron sembrados en medio de cultivo agar centeno. La identificación a nivel de especie se realizó mediante claves morfológicas a nivel microscópico. La investigación fue de tipo experimental y se desarrolló en el laboratorio de microbiología vegetal de la Universidad Nacional Agraria, en Mangua, Nicaragua, bajo un diseño completamente al azar (DCA). Se evaluaron cuatro tratamientos: Metalaxyl + mancozeb, oxicloruro de cobre, Bacillus subtilis, Trichoderma sp., y un testigo absoluto (para determinar el porcentaje de inhibición) con seis repeticiones por tratamiento, totalizando 30 unidades experimentales. Las variables evaluadas fueron identificación de aislados de la rizosfera de papa, capacidad inhibitoria in vitro de Bacillus subtillis, oxicloruro de cobre, metalaxyl+mancozeb, y actividad antagónica de Trichoderma sp. Los resultados obtenidos de capacidad inhibitoria fueron analizados de forma cuantitativa a través de porcentaje de inhibición de crecimiento radial y de forma cualitativa, los datos de antagonismo mediante la escala de cinco clases; se realizó un análisis de varianza para las variables porcentaje de inhibición, mostrando diferencias significativas, siendo el tratamiento más efectivo en capacidad inhibitoria metalaxyl+mancozeb con 88 %, seguido de oxicloruro de cobre con 86 %. Aunque Bacillus subtilis y Trichoderma sp., presentaron bajos porcentajes de inhibición en el crecimiento radial de Phytophthora infestans comparados con los productos sintéticos, estos se clasifican como antifúngico, en el caso de la bacteria Bacillus subtilis y como altamente antagonista el hongo Trichoderma sp.
Palabras clave: antagonismo, patógenos de la papa, Bacillus subtilis, Trichoderma sp., antifúngico, rizosfera.
Abstract: Late blight, caused by the oomycete Phytophthora infestans, is one of the main limiting factors in potato (Solanum tuberosum L.) production in Nicaragua. The aim of this study was to evaluate the in vitro inhibitory capacity of biological and synthetic products for the management of Phytophthora infestans (Late blight). Pathogen isolates were obtained from four farms in the communities of El Robledal and El Zacatón, located in the department of Estelí, northern Nicaragua. Isolation of Phytophthora infestans from soil was carried out using baiting method, employing carnation (Dianthus caryophyllus) leaf fragments as plant bait material; subsequently, these were cultured on rye agar medium. Species-level identification was performed using morphological keys at the microscopic level. The research was experimental and conducted in the plant microbiology laboratory of the Universidad Nacional Agraria, in Managua, Nicaragua, under a completely randomized design (CRD). Four treatments were evaluated: Metalaxyl + mancozeb, copper oxychloride, Bacillus subtilis, Trichoderma sp., and an absolute control (to determine the percentage of inhibition), with six replicates per treatment, totaling 30 experimental units. The evaluated variables included identification of isolates from the potato rhizosphere, in vitro inhibitory capacity of Bacillus subtilis, copper oxychloride, and metalaxyl + mancozeb, as well as antagonistic activity of Trichoderma sp. The results of inhibitory capacity were analyzed quantitatively through percentage inhibition of radial growth and qualitatively using a five-class antagonism data using a five-class antagonism scale. Analysis of variance for inhibition percentage showed significant differences, with the most effective treatment being metalaxyl + mancozeb (88%), followed by copper oxychloride (86%). Although Bacillus subtilis and Trichoderma sp., showed lower percentage of inhibition of radial growth of Phytophthora infestans compared with synthetic products, they are classified as antifungal (in the case of Bacillus subtilis) and highly antagonistic in the case of Trichoderma sp.
Keywords: Antagonism, potato pathogens, Bacillus subtilis, Trichoderma sp., antifungal, rhizosphere.
La papa (Solanum tuberosum L.) es uno de los cultivos de mayor importancia económica y alimentaria a nivel mundial y constituye un rubro estratégico en la agricultura nicaragüense, particularmente en las zonas del norte del país. Sin embargo, su producción se ve severamente afectada por enfermedades de origen fúngico y oomicético, entre las cuales el tizón tardío, causado por Phytophthora infestans, es considerado el principal factor limitante del rendimiento y la estabilidad productiva, debido a su alta agresividad, capacidad de dispersión y potencial destructivo (Miranda y Lanzas, 2007; Pérez y Forbes, 2008).
El manejo del tizón tardío se fundamenta principalmente en el uso preventivo y curativo con fungicidas químicos, como los ditiocarbamatos, compuestos cúpricos y fungicidas sistémicos del grupo de las acilalaninas. Aunque estas herramientas han permitido reducir la severidad de la enfermedad, la aplicación frecuente y repetitiva de ingredientes activos con sitios de acción específicos ha favorecido la selección de poblaciones resistentes de Phytophthora infestans, disminuyendo su eficacia a mediano y largo plazo (Rivera et al., 2014; Robledo, 2015). En Nicaragua, la aparición de aislados resistentes al metalaxyl fue documentada desde la década de 1980, situación que ha persistido debido a la presión de selección ejercida por esquemas de manejo químico poco diversificados (Blandón-Díaz, 2011; Blandón-Díaz et al., 2012).
Además del riesgo de resistencia, el uso intensivo de fungicidas químicos genera efectos adversos sobre el ambiente, la microbiota del suelo y la salud humana, además de incrementar los costos de producción. En este contexto, los programas de manejo integrado de enfermedades promueven la incorporación de estrategias sostenibles que reduzcan la dependencia exclusiva del control químico convencional (Molina et al., 2004; Rivera et al., 2014).
Entre las alternativas emergentes, el control biológico ha mostrado resultados promisorios en el manejo de Phytophthora infestans, destacándose microorganismos antagonistas como Trichoderma spp. y Bacillus subtilis. Estos agentes ejercen su acción mediante diversos mecanismos, como competencia por espacio y nutrientes, antibiosis, producción de metabolitos secundarios y enzimas hidrolíticas, así como la inducción de respuestas defensivas en las plantas hospedantes (Infante et al., 2009; Villareal-Delgado et al., 2018). Estudios previos han demostrado la capacidad inhibitoria in vitro de estos microorganismos frente a Phytophthora infestans, lo que los posiciona como candidatos viables dentro de esquemas de manejo sostenible (García-Núñez et al., 2017; Sánchez, 2016).
Asimismo, fungicidas de acción multisitio, como el oxicloruro de cobre, continúan siendo una herramienta relevante en el manejo del tizón tardío debido a su bajo riesgo de generar resistencia en el patógeno, ya que interfieren en múltiples procesos fisiológicos celulares (Kovačič et al., 2013). No obstante, la efectividad de estos productos puede variar en función de la variabilidad genética de las poblaciones locales de Phytophthora infestans, lo que hace necesaria su evaluación frente a aislados obtenidos en las principales zonas productoras de papa del país.
La evaluación in vitro de la capacidad inhibitoria de productos biológicos y sintéticos frente a Phytophthora infestans constituye una etapa clave para la selección de alternativas de manejo que puedan ser validadas posteriormente en condiciones de campo. En este sentido, el objetivo de la presente investigación fue evaluar la capacidad inhibitoria in vitro de fungicidas químicos y productos biológicos utilizados para el manejo de Phytophthora infestans, como alternativas para el control del tizón tardío en el cultivo de papa (Solanum tuberosum L.).
MATERIALES Y MÉTODOS
Ubicación del área de estudio
El estudio se realizó en el laboratorio de microbiología vegetal de la Universidad Nacional Agraria, ubicada en el km 12 ½ de la Carretera Norte en Managua, capital de Nicaragua, en las coordenadas 12°08'51.05" de latitud Norte y 86°09'50.69" de longitud Oeste.
Diseño experimental
La investigación fue del tipo experimental, de corte transversal y se realizó mediante un diseño completo aleatorizado (DCA). Se implementaron cuatro tratamientos y un testigo absoluto (éste para determinar el porcentaje de inhibición) con seis repeticiones, sumando un total de 30 unidades experimentales (platos Petri). A partir de pruebas in vitro se observó la capacidad inhibitoria de los productos evaluados sobre la base de sus dosis comerciales y dosis expresada en unidades formadoras de colonias de los productos biológicos, las que fueron ajustadas para un menor volumen de agua (Cuadro 1).
Se utilizó la metodología de medio envenenado propuesta por Dingra y Sinclair (1985), técnica que se utiliza para valorar la actividad inhibitoria de microorganismos a nivel de laboratorio (in vitro); luego de inoculadas las placas con los tratamientos se midió el crecimiento radial y la capacidad inhibitoria de los tratamientos sobre el desarrollo de Phytophthora infestans.
Para el método de medio envenenado se utilizó medio de cultivo agar centeno (AC); previo a verter los platos Petri, se adicionaron 10 ml de tratamiento, previamente ajustado a la cantidad a usar, luego se colocó un disco de 5 mm con micelio del patógeno evaluado en el centro del medio, las placas fueron incubadas a 22 °C y se registraron los datos cada 24 horas durante siete días.
| Tratamiento | Descripción | Dosis |
| T0 | Testigo absoluto | Sin aplicación |
| T1 | Trichodermasp | 1x1012 UFC |
| T2 | Bacillus subtilis | 1x108 UFC |
| T3 | Metalaxyl+mancozeb | 0.7 g 100 ml-1 medio agar-centeno |
| T4 | Oxicloruro de cobre | 2 g 100 ml-1 medio agar-centeno |
UFC: Unidades formadoras de colonias.
Colecta de muestras suelo y tejido vegetal
Las muestras de suelo fueron colectadas en tres fincas ubicadas en el departamento de Estelí, en el Norte de Nicaragua. El tipo de muestreo fue simple al azar en forma de “X”, con el objetivo de asegurar una adecuada representatividad de la finca y de la obtención de las muestras (Rios y Baca, 2006). En cada finca se excavaron cinco puntos equidistantes a 50 metros, a una profundidad de 20 cm, con un peso aproximado de 1 kg por muestra.
Para preservar la calidad microbiológica de las muestras y evitar contaminaciones externas, el material fue colocado en bolsas plásticas limpias, previamente rotuladas. Cada etiqueta contenía información que incluía el nombre de la finca, la fecha de muestreo, la profundidad de recolección y las coordenadas geográficas del sitio. La recolección se realizó procurando minimizar la exposición prolongada al sol o a fuentes de calor que pudieran alterar la composición microbiana del suelo. Estas muestras fueron transportadas al laboratorio en un contenedor térmico, donde se procedió a su almacenamiento y posterior aislamiento e identificación del fitopatógeno (Mendoza y Espinoza, 2017).
Para la colecta de hojas se tomó como criterio aquellas que manifestaban síntomas característicos de la enfermedad, que consistían en manchas necróticas con bordes irregulares con halos de coloración amarillenta (Noroña y Tipanquiza, 2010), y se almacenaron en bolsas plásticas con su respectiva etiqueta.
La colecta se realizó en marzo del 2024 en sistemas de producción del cultivo de papa; el primer punto de colecta fue la finca El Robledal ubicada en las coordenadas 13°11'34.5" de latitud Norte y 86°16'57" de longitud Oeste; al momento del muestreo, el cultivo se encontraba en etapa de cosecha. En la segunda finca, Rosita, con coordenadas 13°14'04.6" de latitud Norte y 86°14'29.8" de longitud Oeste; el cultivo tenía aproximadamente 50 días después de la siembra (inicio de tuberización). El último sitio de muestreo se realizó en la finca El Zacatón, con coordenadas 13°13'35.2" de latitud Norte y 86°14'07.8" de longitud Oeste. En la finca Rosita se realizaban aplicaciones de sulfato de cobre pentahidratado (247 g l-1) y otro producto a base de sulfato de cobre pentahidratado (80 %) más sulfato de gentamicina (2 %) más clorhidrato de Oxitetraciclina (6 %) para el manejo de tizón tardío o chamusco como es conocido en la zona; en El Robledal y Zacatón no fueron indicados los métodos de manejo del patógeno.
Aislamiento de Phytophthora infestans a partir de tejido vegetal
Para el aislamiento a partir de hojas de papa se realizaron cortes de 3 mm x 3 mm que abarcara tejido sano y tejido enfermo, posterior a esto se desinfectó con alcohol al 70 % durante 30 segundos, luego se enjuagaron con agua destilada estéril y se dejaron secar en papel toalla, luego se sembraron cinco cortes en platos Petri conteniendo el medio de cultivo agar centeno. Los platos Petri codificados fueron incubados a 22 °C durante 72 horas.
Aislamiento de Phytophthora infestans por método de cultivo trampa
El aislamiento a partir de muestras de suelo se realizó pensando 200 gramos de suelo, luego se depositó en un envase con capacidad de 800 ml, posteriormente se agregaron 500 ml de agua destilada estéril, se homogenizó con una espátula estéril y se dejó reposar hasta que la materia orgánica presente en el suelo se observara sobre la superficie del agua, para posteriormente retirarla con ayuda de papel toalla estéril, seguido de esto se agregaron trozos de hojas de clavel (Dianthus caryophyllus) de tal manera que los propágulos del patógeno se liberaran en el agua, subiendo a la superficie y ocurriendo la infección sobre el tejido vegetal usado como trampa (clavel). Los envases se etiquetaron con los códigos correspondientes y se incubaron durante 72 horas (Soto et al., 2017).
Después de transcurrido las 72 horas, se sembraron en medio de cultivo agar centeno cinco fragmentos de hoja de clavel en un plato Petri, esto se repitió en tres platos Petri por cada aislado, luego las muestras fueron incubadas durante siete días a una temperatura de 22 oC. Para purificar los aislados se hizo uso del método de superposición, consistiendo en colocar un trozo de medio de agar centeno con micelio, colocarlo con el crecimiento hacia arriba y sobre este se colocó un triángulo de medio estéril, posteriormente se incubó por siete días (Soto et al., 2017). La codificación para cada aislado de Phytophthora infestans fue la siguiente: Finca El zacatón (FEZ), Finca Rosita (FRJ) y Finca El Robledal (FER).
Descripción de Phytophthora infestans
Para describir a Phytophthora infestans se utilizaron aislados con buen desarrollo y libres de contaminantes fúngicos y bacterianos, se agregó una gota de lactofenol en un portaobjetos estéril en el que se colocó el micelio y se observó en un microscopio a 40 X. La morfología observada se ingresó en la herramienta interactiva Lucid key (Abad et al., 2023) para la identificación morfológica de Phytophthora, así como el uso de la clave de Gallegly y Hong (2008) y Erwin y Ribeiro (1996). Las características macroscópicas se evaluaron en medio de cultivo jugo V8.
Capacidad inhibitoria in vitro de Bacillus subtilis sobre Phytophthora infestans
Se realizó en medio de cultivo papa dextrosa agar (PDA), similar a la prueba de antagonismo con cultivos duales, siguiendo el procedimiento siguiente: las bacterias fueron obtenidas a partir de un producto comercial y cultivadas en medio agar nutritivo (AN). En un extremo del plato Petri conteniendo papa dextrosa agar, se colocó un disco de 5 mm de cada aislado de Phytophthora infestans, luego se realizó un estriado de la bacteria con la ayuda de un asa bacteriológica previamente esterilizada en línea recta centrada (Oliva et al., 2017) (Figura 1) y se incubó a 22 °C; el crecimiento micelial se midió en dirección al centro del plato Petri cada 24 horas y con los datos obtenidos se realizó el cálculo de porcentaje de inhibición.
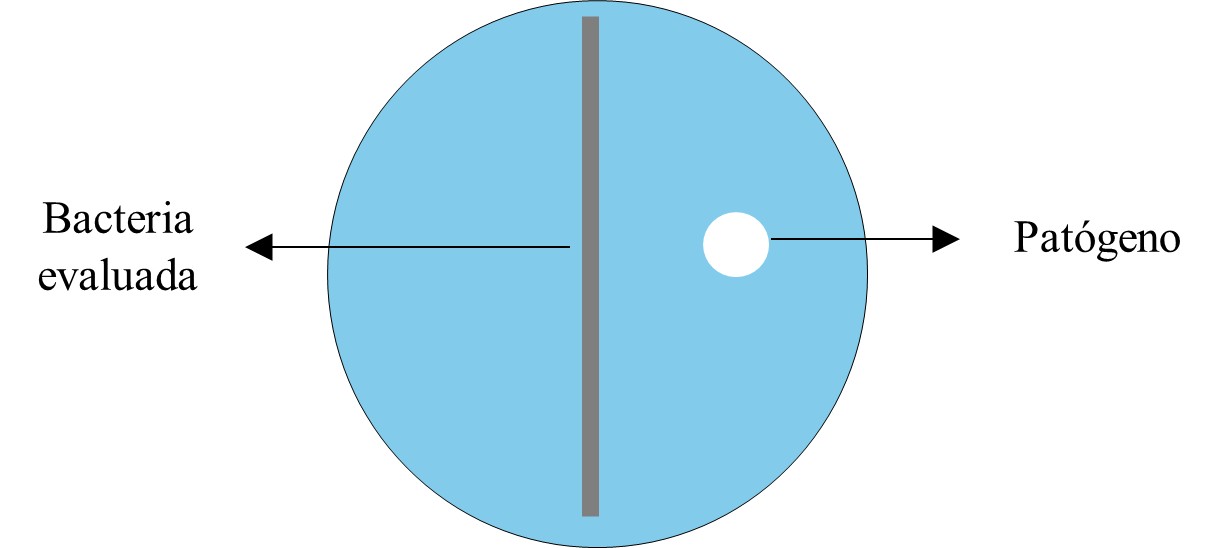
Figura 1.
Representación de prueba de inhibición de B. subtilis sobre P. infestans.
Inhibición de crecimiento micelial de Phytophthora infestans con oxicloruro de cobre y metalaxyl + mancozeb
Se realizó en medio de cultivo agar centeno (AC) mediante la técnica de medio envenenado propuesta por Dingra y Sinclair (1985), que consiste en adicionar 10 ml del tratamiento a evaluar en el medio de cultivo agar centeno (AC), luego se vertieron en platos Petris y se dejaron solidificar; una vez solidificado el medio de cultivo, se colocó un disco de 5 mm con micelio de Phytophthora infestans, las placas fueron incubadas a 22 °C y se registraron los datos cada 24 horas, midiendo el crecimiento radial a partir del centro del disco hacia los extremos.
Actividad antagónica de Trichoderma sp., sobre Phytophthora infestans
La actividad antagónica se evaluó mediante la técnica de cultivo dual in vitro, con medio PDA vertido en platos Petri, y se inocularon discos con micelio de 5 mm de ambos microorganismos en extremos opuestos del plato Petri. Los platos Petri se incubaron a 24 °C durante siete días, monitoreando de forma visual, realizando medición de crecimiento radial de los micelios, así como la evaluación de las zonas de inhibición, la interacción entre ambos micelios y comparación con el testigo absoluto, todo con intervalos de 12 horas; con una regla milimitrada se midió desde el centro del disco del antagonista en dirección al disco con micelio de Phytophthora infestans, hasta que el testigo absoluto cubrió toda el área del plato Petri conteniendo el medio de cultivo, esto permitió determinar la capacidad inhibitoria de Trichoderma sobre Phytophthora infestans de forma cualitativa y cuantitativa (Cuadro 2), según lo descrito por Bell et al. (1982). El porcentaje de inhibición del antagonista sobre Phytophthora infestans se calculó mediante la fórmula propuesta por Ezziyyani et al. (2004):
I (%) = [(Tc - Tt) / Tc] x 100
I (%): Porcentaje de inhibición
Tc: Radio promedio del testigo absoluto (T0)
Tt: Radio de la colonia en medio con el antagonista
Evaluación del grado de antagonismo
Se midió el grado de antagonismo con el uso de la escala propuesta por Bell et al. (1982), dicha escala cuenta con cinco clases, indicando que Trichoderma sp., es altamente antagonista si alcanza un valor promedio igual o menor a 2 y no es considerado altamente antagonista si el valor promedio es igual o mayor a 3.
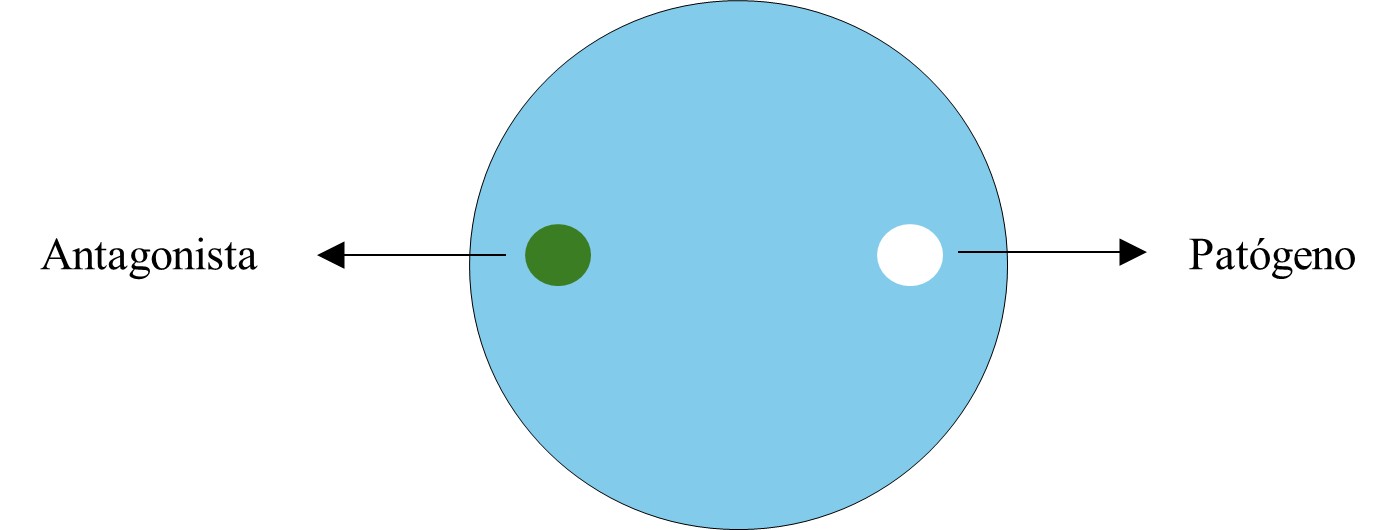
Figura 2.
Representación de cultivos duales para prueba de antagonismo
| Clase | Descripción |
| 1 | El antagonista crece completamente sobre la colonia del patógeno y cubre toda la superficie del medio de cultivo |
| 2 | El antagonista cubre al menos las dos terceras partes de la superficie del medio de cultivo |
| 3 | El antagonista y el patógeno cubren aproximadamente la mitad de la superficie del medio de cultivo |
| 4 | El patógeno cubre al menos dos terceras partes del medio de cultivo limitando el crecimiento del antagonista |
| 5 | El patógeno crece sobre la colonia del antagonista y cubre toda la superficie del medio de cultivo |
Fuente: Bell et al. (1982).
Colecta de datos
Se elaboró una base de datos en Microsoft Excel (2016) en la que se registró información de las mediciones por tratamiento según intervalos de 12 horas o 24 horas de medición a partir de la inoculación en los platos Petri. El registro de datos para el porcentaje de crecimiento radial se realizó hasta cumplir las 96 horas y el grado antagónico hasta cumplir 144 horas.
Variables evaluadas
Las variables evaluadas consistieron en la descripción de aislados de Phytophthora infestans a partir de muestras del cultivo de papa (rizosfera), la capacidad inhibitoria in vitro de Bacillus subtilis, Metalaxyl+Mancozeb y oxicloruro de cobre, sobre Phytophthorainfestans en dosis comerciales, así como la actividad antagónica y el grado de antagonismo de Trichoderma sp. El porcentaje de inhibición in vitro de Bacillus subtilis, metalaxyl+mancozeb y oxicloruro de cobre sobre Phytophthora infestans en dosis comerciales se calculó con la fórmula para el cálculo de porcentaje de inhibición.
Análisis de datos
Las variables porcentaje de inhibición de Bacillus subtilis, metalaxyl+mancozeb, oxicloruro de cobre y Trichoderma sp se evaluaron de forma cuantitativa, obteniendo porcentaje de inhibición de crecimiento radial. La variable grado antagónico de Trichoderma sp., fue analizada de forma cualitativa mediante la escala de 5 clases (Cuadro 2). Las variables porcentaje de inhibición y grado antagónico se sometieron a una prueba de normalidad, al cumplirse el supuesto de normalidad se procedió a realizar un análisis de varianza (ANDEVA) y separación de medias mediante la prueba de Tuckey α 0.05 con el uso del Software estadístico InfoStat versión libre (2021).
RESULTADOS Y DISCUSIÓN
Descripción de Phytophthora infestans aisladas a partir de la rizosfera del cultivo de papa
Para la identificación de Phytophthora infestans a nivel de especie, se consideró la morfología de las características macroscópicas y microscópicas, como el tipo de crecimiento micelial, que presentó un crecimiento micelial aéreo abundante, con coloración blanca, además, presentó una textura algodonosa y con crecimiento de micelio en forma de roseta en el medio de cultivo jugo V8 característico de Phytophthora infestans (Figura 3). Se observó que los aislados fueron heterotálicos, con estructuras reproductivas asexuales como esporangios caducos, semipapilados y limoniformes, así como hifas cenocíticas no abultadas (Figura 4), además se identificaron estructuras reproductivas sexuales como es el caso de oogonios de paredes lisas con la unión del anteridio anfígeno, y oosporas pleróticas (Figura 5).

Figura 3.
Colonia de Phytophthora infestans en medio V8 con características de crecimiento roseteado
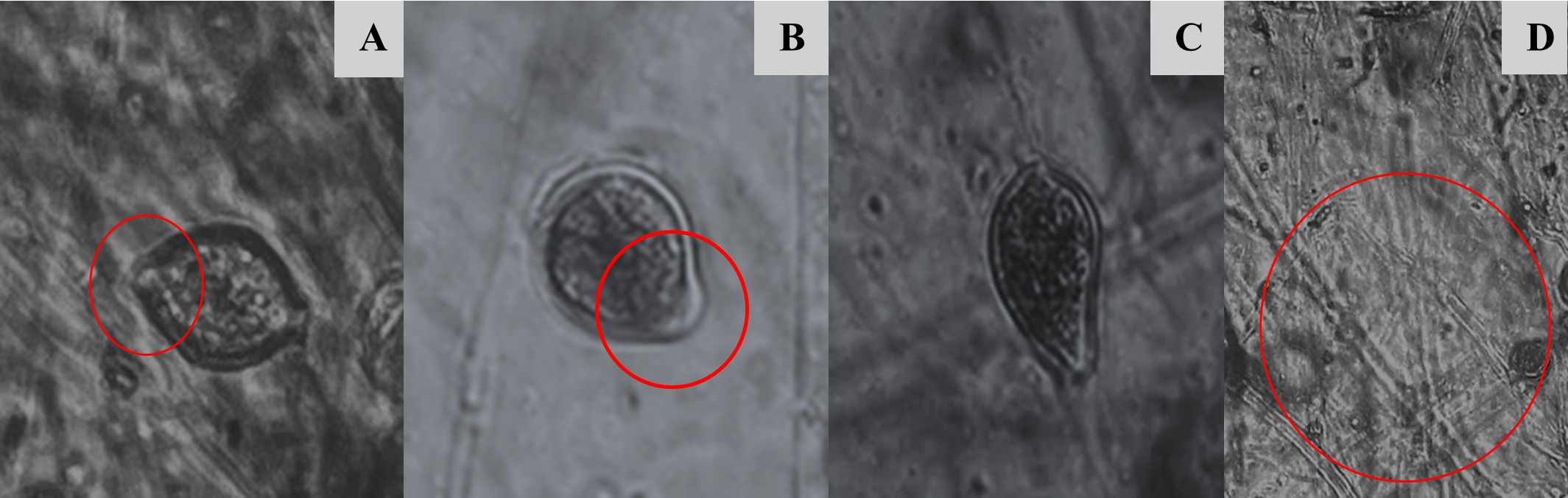
Figura 4.
Estructuras reproductivas asexuales de Phytophthora infestans; A y B: esporangios semipapilados, C: esporangio limoniforme y D: Hifas no abultadas.
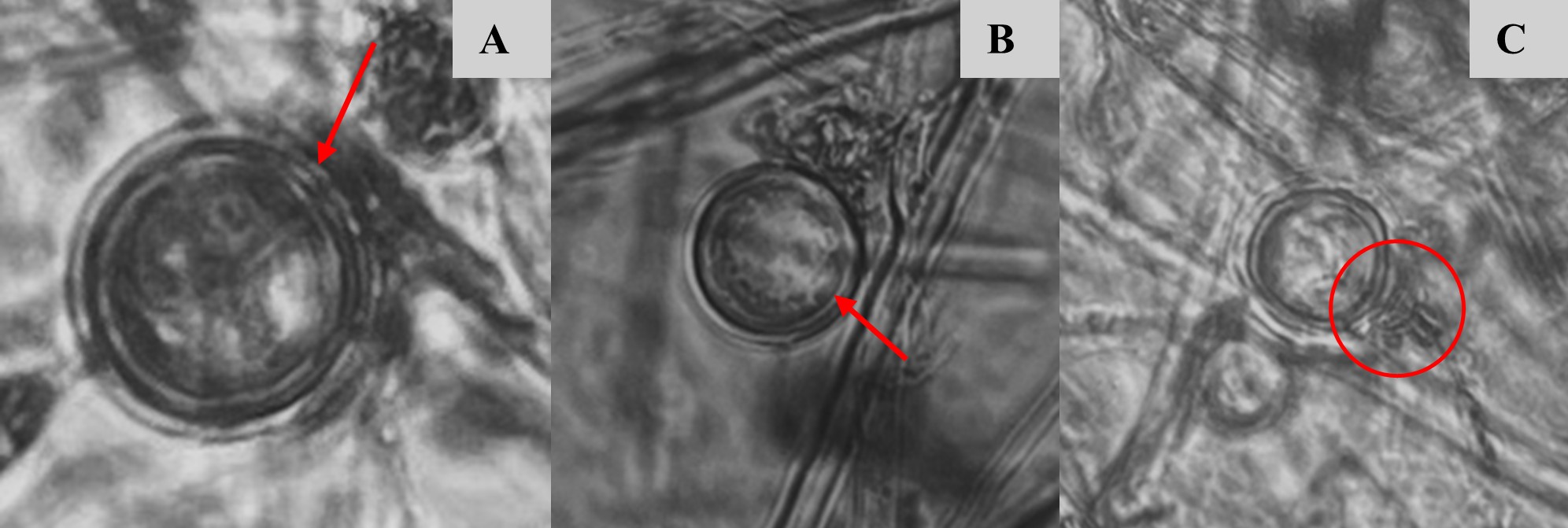
Figura 5.
Estructuras reproductivas sexuales de Phytophthora infestans ; A y B: oogonios con oospora plerótica y C: oogonio con unión del anteridio anfígeno.
La identificación a nivel microscópico coincide con las características descritas por Erwin y Ribeiro (1996), Gallegly y Hong (2008) y los resultados emitidos por la herramienta interactiva IDphy (Lucid key), donde indican que Phytophthora infestans presenta esporangios semipapilados, y caducos, desprendiéndose fácilmente del esporangióforo, además de que esta especie está caracterizada por ser heterotálica, a diferencia de otras especies de Phytophthora que son homotálicas.
Inhibición de crecimiento radial de Bacillus subtilis sobre Phytophthora infestans
El porcentaje de inhibición del crecimiento radial (PICR) de Bacillus subtilis sobre Phytophthora infestans para los aislados de la fina El Zacatón y la finca El Robledal los resultados fueron igual o mayor de 70 % siendo inferior el obtenido del aislado de la finca Rosita (Figura 6).
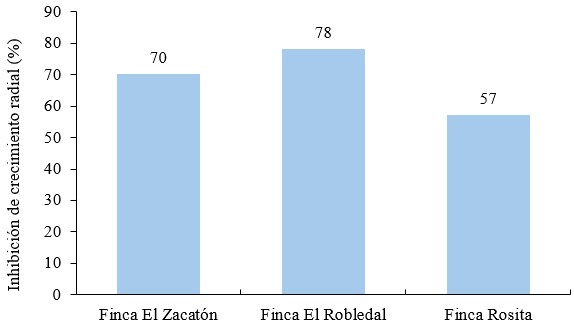
Figura 6.
Inhibición de crecimiento radial (%) de Bacillus subtillis en aislados de Phytophthora infestans según sistema de producción
Los porcentajes obtenidos en los aislados FEZ y FER fueron de 70 % y 78 % respectivamente (Figura 7), encontrándose en un rango similar a los obtenidos por Wang et al. (2020) donde la cepa Bacillus subtilis inhibió en un 75.6 %, lo que representa un mejor control sobre Phytophthora infestans en comparación con Pseudomonas sp y Bacillus pumillus; en cambio, Alfiky et al. (2022) obtuvieron un porcentaje de 82 % con una cepa de Bacillus subtilis.
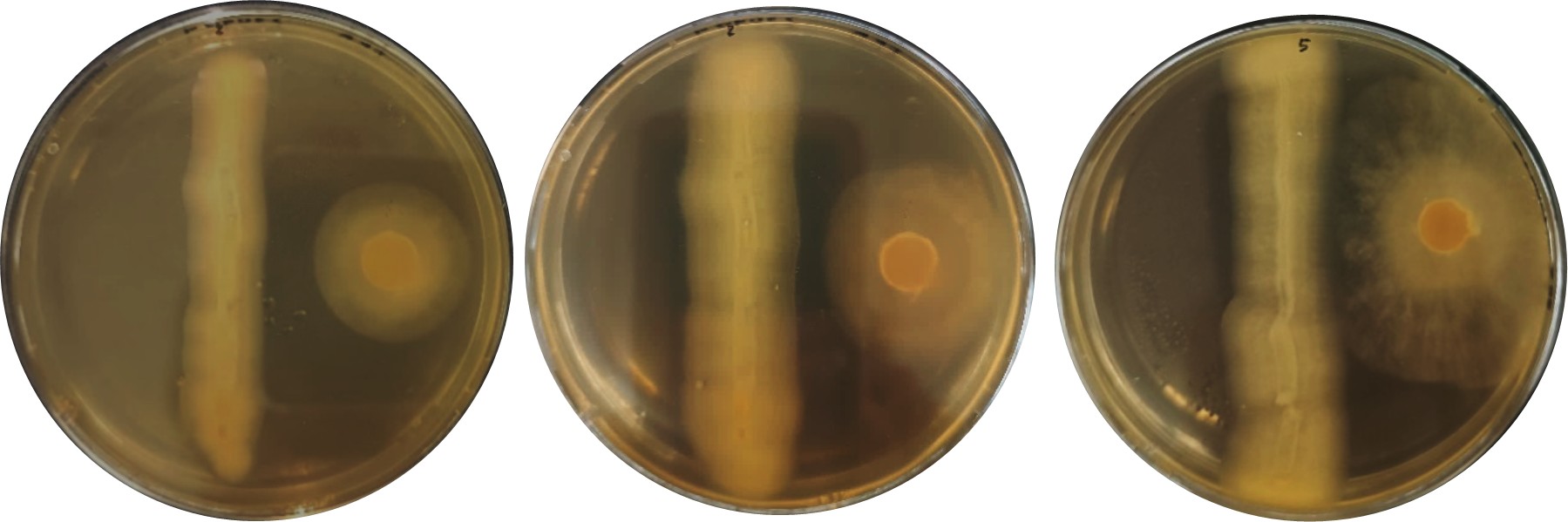
Figura 7.
Inhibición de Bacillus subtilis sobre aislados de Phytophthora infestans; A: FER, B: FEZ y C: FRJ.
Villareal-Delgado et al. (2018), mencionan que la capacidad antifúngica de Bacillus subtilis se basa en la producción de moléculas de naturaleza química como son los lipopéptidos (surfactina, iturina y fengicina), que inhiben la expresión de un microorganismo patógeno, afectando la membrana citoplasmática de las células fúngicas, y como consecuencia generan la muerte celular.
La especie Bacillus subtilis es una alternativa eficaz para prevenir el desarrollo de Phytophthora infestans, tal como lo menciona Arteaga (2023), quien indica que la actividad biológica inhibitoria de Bacillus subtilis sobre Phytophthora sp., in vitro e in vivo, reducen significativamente el crecimiento micelial del patógeno, con valores de inhibición mayores al 70 %.
Inhibición de crecimiento radial de metalaxyl + mancozeb sobre Phytophthora infestans
El porcentaje de inhibición de crecimiento radial obtenido en los aislados provenientes de las tres fincas (FEZ, FER y FRJ) fueron mayores al 85 %, por tanto, son considerados como sensibles al efecto inhibitorio ejercido por el tratamiento (Figura 8).
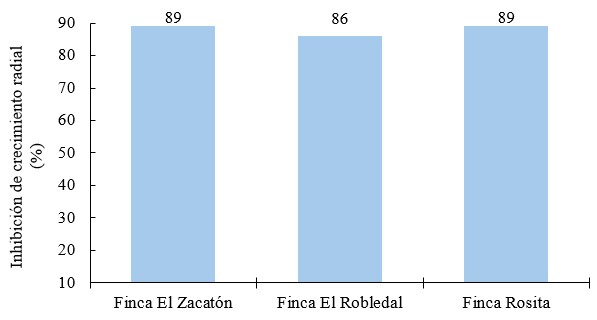
Figura 8.
Inhibición (%) de metalaxyl+mancozeb en aislados de Phytophthora infestans
Los criterios para clasificar los aislados de Phytophthora infestans como sensibles, intermedios o resistentes al metalaxyl se basan en el crecimiento radial de la colonia en presencia del fungicida, comparado con los tratamientos testigos (Riveros et al., 2003). Considerando que el diámetro total del plato Petri es de 90 mm, el porcentaje de inhibición se calcula en función del crecimiento radial observado. Los aislados se clasifican según Riveros et al. (2003) como sensibles si presentan un porcentaje de crecimiento radial menor al 10 % (crecimiento inferior a 9 mm), intermedios con un porcentaje de crecimiento radial entre el 10 % y el 60 % (crecimiento entre 9 mm y 54 mm), y resistentes si el porcentaje de crecimiento radial es igual o mayor al 60 % (crecimiento superior a 54 mm).
Páez et al. (2001) mencionan que los ingredientes activos pertenecientes al grupo químico de las acilalalinas, tal es el caso de metalaxyl, actúan sobre sitios específicos del patógeno, debido a esta característica se encontró en la evaluación de resistencia in vitro e in vivo de Phytophthora infestans al metalaxyl, que de 69 aislados en estudio, el 64 % (44 aislados), presentaron resistencia y un 36 % (25 aislados), fueron sensibles.
El modo de acción de este ingrediente activo es inhibir la enzima RNA polimerasa y ARN ribosomal, inhibiendo así la síntesis de proteínas y reduciendo el desarrollo micelial, sin embargo, al ser Phytophthora infestans un microorganismo considerado con alta variabilidad genética, genera resistencia dado los cortos periodos de aplicación del producto (Robledo, 2015).
En Nicaragua, la resistencia y el aumento de aislados de Phytophthora infestans al metalaxyl ocurrieron en la década de 1980 hasta 1990, debido que los productores de papa comenzaron a utilizar este ingrediente activo de manera indiscriminada y frecuente. Consecuente a esto, se condujo a una presión de selección por parte de aislados de Phytophthora infestans, donde persiste hasta la actualidad en la población clonal nicaragüense de este patógeno (Blandón-Díaz, 2011).
Para evitar la aparición de más poblaciones del patógeno resistente se comenzó a aplicar con otros ingredientes activos, tal es el caso de la combinación metalaxyl+mancozeb que se empezó a utilizar a partir del año 1985 (Robledo, 2015), combinación utilizada en este estudio, en el que se evidencia que, al utilizar este ingrediente activo con otro de diferente modo de acción, sigue siendo eficaz para disminuir el desarrollo del patógeno.
Inhibición de crecimiento radial de oxicloruro de cobre sobre P. infestans
El porcentaje de inhibición del crecimiento radial (PICR) de oxicloruro de cobre sobre Phytophthora infestans en todos los aislados mostraron resultados mayores al 80 % (Figura 9). No se observó crecimiento radial del microorganismo desde el inicio hasta el final del periodo de incubación.
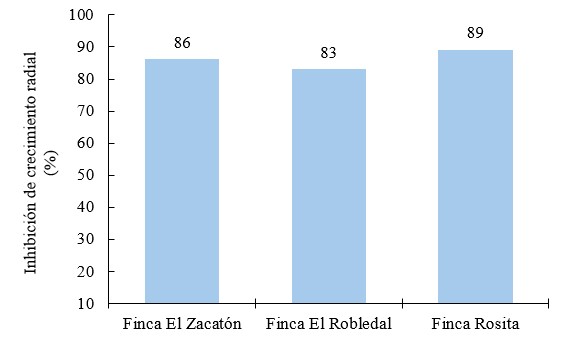
Figura 9.
Inhibición de crecimiento radial (%) de oxicloruro de cobre sobre aislados Phytophthora infestans
Estudios realizados por Gopi et al. (2020) a nivel de campo en el cultivo de tomate en el que se evaluaron extractos de ajo (Allium sativum), Trichoderma sp., hidróxido de cobre y oxicloruro de cobre, este último presentó 87.52 % de control sobre el crecimiento de la infección.
El riesgo de que un producto a base de cobre genere resistencia es bajo, dado que en el patógeno se bloquean las enzimas relacionadas al proceso de respiración, se desnaturalizan las proteínas y también provoca envenenamiento y muerte celular (Kovačič et al., 2013).
Actividad antagónica de Trichoderma sp., sobre P. infestans
El porcentaje de inhibición del crecimiento radial (PICR) de Trichoderma sp., sobre los aislados de P. infestans en las tres fincas fueron superior a 30 % pero inferiores a 50 % (Figura 10).
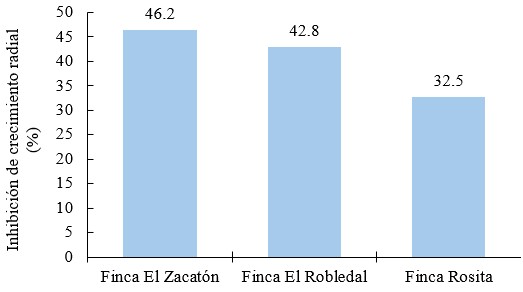
Figura 10.
Inhibición de crecimiento radial (%) a las 144 horas después de la incubación de P. infestans, con aislados de Trichoderma sp
El PICR del aislado de la finca Rosita presentó una menor susceptibilidad a Trichoderma sp, alcanzando valores similares a los reportados por Debebe et al. (2011), donde el valor mínimo de biocontrol de Trichoderma sobre Phytophthora infestans fue de 36.7 %, siendo un resultado cercano al obtenido en esta investigación. Esto a su vez coincide con los resultados obtenidos por Kariuki et al. (2020), quienes indican que el efecto antagónico de Trichoderma harzianum y Trichoderma asperellum sobre Phytophthora infestans, redujeron significativamente el crecimiento micelial del patógeno, siendo la especie de Trichoderma asperellum la que tuvo valores similares al registrado en este estudio con un PICR igual a 30.7 % a los ocho días después del enfrentamiento dual (192 horas después de la incubación).
El asilado de la finca El Zacatón fue más susceptible al efecto antagónico ejercido por la cepa de Trichoderma sp., obteniendo valores similares de inhibición a los reportados por García-Núñez et al. (2017), quienes registraron valores de PICR de 49 % con Trichoderma asperellum sobre Phytophthora infestans. Michel-Aceves et al. (2009), reportan especies de Trichoderma sp., con un porcentaje de biocontrol in vitro entre 5.35 % y 62.9 %.
Sin embargo, en la observación directa de los enfrentamientos en cultivos duales se determinó el efecto antagónico ejercido por Trichoderma sp., sobre el crecimiento micelial de los tres aislados de Phytophthora infestans y presentaron un crecimiento acelerado de Trichoderma en comparación de los aislados de Phytophthora infestans, disminuyendo su desarrollo debido a la competencia por espacio y nutrientes ejercida por el antagonista.
De Aguiar et al. (2014), manifiesta que a pesar de que el aislado de Trichoderma sp., presentó diferentes PICR sobre los aislados de Phytophthora infestans, sigue siendo una buena alternativa de biocontrol, debido a que se caracteriza a Phytophthora infestans como un patógeno virulento.
Evaluación del grado de antagonismo de Trichoderma
Un aislado de Trichoderma se considera altamente antagonista si el valor promedio de la escala es menor o igual a 2 y no es considerado altamente antagonista si el valor promedio es mayor o igual a 3 (Bell et al., 1982); en esta investigación el aislado de Trichoderma sp., se ubicó en la clase 1 de la escala, por lo que es considerado como altamente antagonista sobre los aislados de Phytophthora infestans (Figura 11), lo que lo convierte en un agente viable para poder ser implementado en estudios posteriores a nivel de campo.
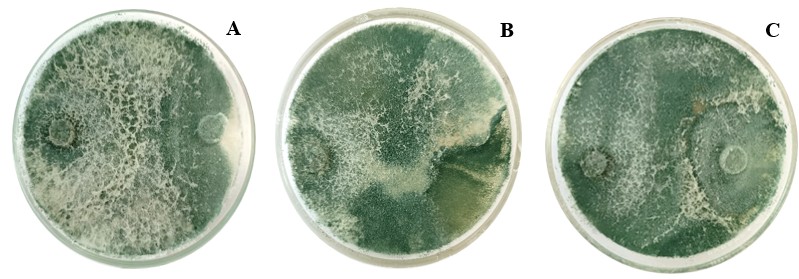
Figura 11.
Antagonismo por competencia ejercido por Trichoderma sobre aislados de Phytophthora infestans; A: finca el Robledal, B: finca El Zacatón y C: finca Rosita.
En estudios realizados sobre la caracterización del agente causal de la pudrición radicular del aguacate, y Trichoderma sp., como potencial agente de control biológico, se menciona lo siguiente: El establecer la capacidad antagónica de las cepas de Trichoderma en condiciones in vitro es algo necesario y tiene como principio la selección de las mejores cepas, esto para las pruebas posteriores a nivel de campo, además que se genera información sobre una posible formulación con materiales inertes apropiados que le brinden mayor efectividad; esto permite obtener resultados satisfactorios en el manejo biológico en los agroecosistemas donde los fitopatógenos puedan repercutir de manera negativa (Sánchez, 2016).
En este estudio se observó que la capacidad antagónica de Trichoderma sp., fue atribuida a uno de sus mecanismos de acción, como es el caso de competencia por espacio y nutrientes (Figura 11). Infante et al. (2009) indican que Trichoderma sp., al ser un microorganismo de crecimiento acelerado y rápido desarrollo, el principal mecanismo de acción es la competencia por espacio y nutrientes con otros microorganismos que se encuentran en el mismo sustrato. Además, se realizan numerosos estudios de especies que pertenecen al género Trichoderma para ser evaluados y propuestos como una alternativa de control biológico debido a sus características de crecimiento rápido en una alta gama de sustratos, producción de enzimas (quitinasas, glucanasas, proteasas) y metabolitos secundarios que degradan la pared celular de microorganismos fitopatógenos, además de tener resistencia a diversos fungicidas (Tyśkiewicz et al., 2022).
Inhibición de crecimiento radial por tratamiento frente a Phytophthora infestans
Existen diferencias estadísticas entre el porcentaje de inhibición del crecimiento radial (p = 0.0004). Los mejores tratamientos fueron metalaxyl+mancozeb (89 %) y oxicloruro de cobre con 86 %; Bacillus subtilis mostró una efectividad al inhibir en un 68 % y Trichoderma sp., en 40.6 %. A pesar de que el porcentaje de inhibición de los productos no biológicos superan a los biológicos, se destaca que Trichoderma sp., se presenta como un hongo altamente antagonista a Phytophthora infestans y Bacillus subtilis como una bacteria con alta capacidad antifúngica.
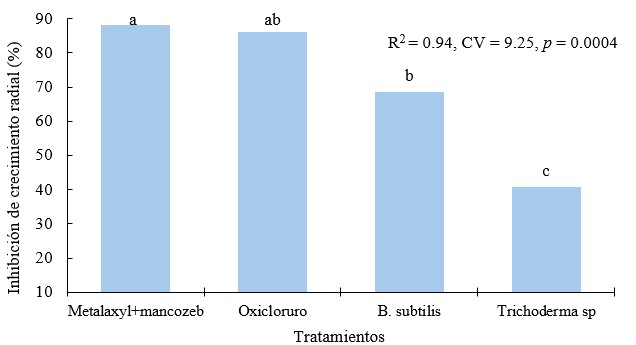
Figura 12.
Inhibición de crecimiento radial por tratamiento sobre aislados de Phytophthora infestans
CONCLUSIÓN
Se presentan distintos niveles de eficacia in vitro frente a Phytophthora infestans, confirmando la viabilidad tanto de fungicidas sintéticos como de alternativas biológicas para el manejo de Phytophthora infestans, agente causal del tizón tardío. El producto biológico a base de Bacillussubtilis mostro ser efectivo con 68 % de inhibición radial, y Trichoderma sp., como altamente antagonista. Los productos biológicos pueden ser una alternativa para el manejo en campo de este patógeno.
REFERENCIAS BIBLIOGRÁFICAS
Abad Z., G., Burgess, T. I., Redford A., J, Bienapfl J., C., Coffea, M., & Caballero, L. (2023). IDphy: Molecular and morphological identification of Phytophthora based on the types. USDA APHIS PPQ S&T Plant Pathogen Confirmatory Diagnostics Lab; USDA APHIS PPQ S&T ITP; Phytophthora Science and Management, Murdoch University; World Phytophthora Collection. https://idtools.org/phytophthora/index.cfm?pageID=3141
Alfiky, A., L’Haridon, F., Abou-Mansour, E & Weisskopf, L. (2022). Disease inhibiting effect of strain Bacillus subtilis EG21 and its metabolites against potato pathogens Phytophthora infestans and Rhizoctonia solani. Phytopathology, 112(10), 2099-2109. https://doi.org/10.1094/PHYTO-12-21-0530-R
Arteaga, R. P. (2023). Evaluación de la actividad biológica inhibitoria de filtrados producidos por cultivos inmovilizados en agar-agar de Bacillus sp. contra Phytophthora sp. como patógeno foliar del tomate (Solanum lycopersicum) [Tesis de licenciatura, Universidad Mayor de San Andrés]. https://repositorio.umsa.bo/bitstream/handle/123456789/35259/TG-2053.pdf?sequence=1&isAllowed=y
Bell, D. K., Wells, H. D., & Markham, C. R. (1982). In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology, 72(4), 379-382. https://doi.org/10.1094/Phyto-72-379
Blandón-Díaz, J. U. (2011). Insights into population structure and epidemiology of Phytophthora infestans from Nicaragua. Department of Forest Mycology and Pathology, Swedish University of Agricultural Sciences. https://core.ac.uk/reader/11697700
Blandón-Díaz, J. U., Widmark, A. K., Hannukkala, A., Andersson, B., Högberg, N., & Yuen, J.E. (2012). Phenotypic variation within a clonal lineage of Phytophthora infestans infecting both tomato and potato in Nicaragua. Phytopathology, 102(3), 323-330. https://doi.org/10.1094/PHYTO-02-11-0033
De Aguiar, R. A., da Cunha, M. G., & Junior, M. L. (2014). Management of white mold in processing tomatoes by Trichoderma spp. and chemical fungicides applied by drip irrigation. Biological Control, 74, 1-5. https://doi.org/10.1016/j.biocontrol.2014.03.009
Debebe, Z, E., Santhanam, A., Gorfu, D., Tessera, M., & Kassa, B. (2011). Biocontrol activity of Trichoderma viride and Pseudomonas fluorescens against Phytophthora infestans under greenhouse conditions. Journal of Agricultural Technology, 7(6), 1589-1602. http://www.ijataatsea.com/pdf/v7_n6_11_November/12_IJAT_2011_7_6___Ephrem_Debebe_Zegeye-22%20Nov%202011.pdf
Di Rienzo, J. A., Casanoves, F., González, L. A., Tablada, M. G., Robledo, C. W., & Balzarini, M. G. (2021). InfoStat (Versión 2021) [Software]. Grupo InfoStat. https://infostat.com.ar
Dingra, D., & J. Sinclair. (1985). Basic Plant Pathology Methods. Editorial CRC Press.
Erwin, D. C., & Ribeiro, O. K. (1996). Phytophthora Diseases Worldwide. APS Press.
Ezziyyani, M., Pérez, C., Requena, M. E., Rubio, L., & Candela, M. E. (2004). Biocontrol por Streptomyces rochei –Ziyani–, de la podredumbre del pimiento (Capsicum annuum L.) causada por Phytophthora capsici. Anales de Biología, 26, 69-78. https://revistas.um.es/analesbio/article/view/30471
Gallegly, M. E., & Hong, C. (2008). Phytophthora: Identifying species by morphology and DNA fingerprints. The American Phytopathological Society.
García-Núñez, H. G., Martínez-Campos, Á. R., Hermosa-Prieto, M. R., Monte-Vázquez, E., Aguilar-Ortigoza, C. J., & González-Esquivel, C. E. (2017). Morphological and molecular characterization of native isolates of Trichoderma and its potential biocontrol against Phytophthora infestans. Revista mexicana de fitopatología, 35(1), 58-79. https://doi.org/10.18781/r.mex.fit.1605-4
Gopi, R., Avasthe, R. K., Kalita, H., Yadav, A., Das, S. K., & RAI, D. (2020). Eco-friendly management of tomato late blight using botanicals, bio-control agents, compost tea and copper fungicides. The Indian Journal of Agricultural Sciences, 90(1), 35-39. https://doi.org/10.56093/ijas.v90i1.98525
Infante, D., Martínez, B., González, N., y Reyes, Y. (2009). Mecanismos de acción de Trichoderma frente a hongos fitopatógenos. Revista de protección vegetal, 24(1), 14-21.https://www.researchgate.net/publication/262498589_mecanismos_de_accion_de_trichoderma_frente_a_hongos_fitopatogenos
Kariuki, W. G., Mungai, N. W., Otaye, D. O., Thuita, M., Muema, E., Korir, H., & Masso, C. (2020). Antagonistic effects of biocontrol agents against Phytophthora infestans and growth stimulation in tomatoes. African Crop Science Journal, 28(s1), 55-70. https://doi.org/10.4314/acsj.v28i1.5S
Kovačič, G. R., Lešnik, M., & Vršič, S. (2013). An overview of the copper situation and usage in viticulture. Bulgarian Journal of Agricultural Science, 19(1), 50-59. https://www.agrojournal.org/19/01-07.pdf
Mendoza, R. B. y Espinoza, A. (2017). Guía técnica para muestreos de suelo. https://repositorio.una.edu.ni/3613/1/P33M539.pdf
Michel-Aceves, A. C., Otero-Sánchez, M. A., Solano-Pascacio, L. Y., Ariza-Flores, R., Barrios-Ayala, A., & Rebolledo-Martínez, A. (2009). Biocontrol in vitro con Trichoderma spp. de Fusarium subglutinans (Wollenweb. y Reinking) Nelson, Toussoun y Marasas y F. oxysporum Schlecht., Agentes Causales de la" Escoba de Bruja" del Mango (Mangifera indica L.). Revista mexicana de fitopatología, 27(1), 18-26. https://www.redalyc.org/pdf/612/61211414003.pdf
Miranda, Y. U. y Lanzas, W, J. (2007). Evaluación de alternativas químicas y biológicas en el manejo del tizón tardío [Phytophthora infestans (Mont.) De Bary] en el cultivo de papa (Solanum tuberosum) [Tesis de ingeniería, Universidad Nacional Agraria]. Repositorio Institucional. https://repositorio.una.edu.ni/2024/1/tnh20m672.pdf
Molina, J. D. D., Mairena Santos, B. y Aguilar, B. L. (2004). Manejo integrado de plagas: A MIP en el cultivo de la papa. Instituto Nicaragüense de Tecnología Agropecuaria. https://cenida.una.edu.ni/relectronicos/RENH10M722.pdf
Noroña, C. J. P. y Tipanquiza, I. J. J. (2010). Evaluación del comportamiento en el cultivo de papa (solanum tuberosum) variedades" INIAP; Natividad, INIAP Estela"; versus un testigo, la variedad tradicional" Bolona" en el cantón Paute provincia del Azuay [Tesis de ingeniería, Universidad Politécnica Salesiana]. Repositorio Institucional. http://dspace.ups.edu.ec/handle/123456789/4744
Oliva-Ortiz, L., Velázquez-Alcaraz, T., Sosa-Pérez, R., Ruvalcaba, L., Valdés, T., Arciniega-Ramos, J., & López Orona, C. (2017). Controlling Fusarium wilt of chickpea (Cicer arietinum L.) with native microorganisms of Sinaloa, Mexico. Agrociencia,51(6), 683-695. https://www.researchgate.net/publication/320145714_Controlling_Fusarium_wilt_of_chickpea_Cicer_arietinum_L_with_native_microorganisms_of_Sinaloa_Mexico
Páez, O., Gómez, L., Brenes, A. y Valverde, R. (2001). Resistencia de aislamientos de Phytophthora infestans al metalaxyl en el cultivo de la papa en Costa Rica. Agronomía Costarricense, 25(1), 33-44. http://www.redalyc.org/articulo.oa?id=43625103
Pérez, W. y Forbes, G. (2008). El tizón tardío de la papa. Centro Internacional de la Papa (CIP). https://cipotato.org/wp-content/uploads/2014/08/004271.pdf
Rios, F. y Baca, P. (2006). Niveles y Umbrales de Daños Económicos de las Plagas. Zamorano. https://bdigital.zamorano.edu/server/api/core/bitstreams/16ecca50-98ca-4116-be3a-933d5a388603/content
Rivera, J. M., Brown, J. K., Weller, S. y Melgar, J. C. (2014). Consideraciones técnicas para el efectivo manejo integrado del tizón tardío en papa. La Lima, Cortes, Honduras.http://bvirtual.infoagro.hn/xmlui/bitstream/handle/123456789/180/consideraciones_tecnicas_para_manejo_de_tizon_tardio_en_papa.pdf?sequence=1
Riveros, F., Sotomayor, R., Rivera, V. y Espinoza, B. (2003). Resistencia de Phytophthora infestans (Montagne) de Bary a Metalaxyl, en cultivo de papas en el norte de Chile. Agricultura Técnica, 63(2), 117-124. https://hdl.handle.net/20.500.14001/36845
Robledo, M. N. (2015). Susceptibilidad/resistencia de Phytophthora infestans (Mont) de Bary a fungicidas [Tesis de doctoral, Instituto de enseñanza e investigación en ciencias agrícolas]. http://colposdigital.colpos.mx:8080/xmlui/bitstream/handle/10521/2510/Robledo_Esqueda_MN_DC_Fitopatologia_2014.pdf?sequence=1&isAllowed=y
Sánchez, I. E. (2016). Caracterización del agente causal de la pudrición radicular del aguacate y Trichoderma sp., como potencial agente de control biológico en Nicaragua [Tesis de maestría, Universidad Nacional Agraria]. Repositorio Institucional. https://repositorio.una.edu.ni/3359/1/tnh20s194.pdf
Soto, A., Rodríguez, G., Fernández, Y. L., Pedraza, M. E., López, L., Díaz, M. y Fernández, S. P. (2017). Protocolos de aislamiento y diagnóstico de Phytophthora spp. enfoque aplicado a la investigación. Revista mexicana de ciencias agrícolas, 8(8), 1867-1880. https://doi.org/10.29312/remexca.v8i8.708
Tyśkiewicz, R., Nowak, A., Ozimek, E., & Jaroszuk-Ściseł, J. (2022). Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. International Journal of Molecular Sciences, 23(4), 2329. https://doi.org/10.3390/ijms23042329
Villarreal-Delgado, M. F., Villa-Rodríguez, E. D., Cira-Chávez, L. A., Estrada-Alvarado, M. I., Parra-Cota, F. I. y Santos-Villalobos, S. D. L. (2018). El género Bacillus como agente de control biológico y sus implicaciones en la bioseguridad agrícola. Revista mexicana de fitopatología, 36(1), 95-130. https://doi.org/10.18781/r.mex.fit.1706-5
Wan, Y., Zhang, C., Liang, J., Wu, L., Gao, W., & Jiang J. (2020). Iturin A extracted from Bacillus subtilis WL-2 affects Phytophthora infestans via cell structure disruption, oxidative stress, and energy supply dysfunction. Frontiers in microbiology, 11. https://doi.org/10.3389/fmicb.2020.536083
Notas de autor
Información adicional
redalyc-journal-id: 7858