Ciencia de las Plantas
Respuesta de poblaciones silvestres de tomate (Solanum lycopersicum L.) a la bacteria Ralstonia solanacearum en las etapas de germinación y plántula
Response of wild populations of Solanum lycopersicum L. to Ralstonia solanacearum during germination and seedling stages


Respuesta de poblaciones silvestres de tomate (Solanum lycopersicum L.) a la bacteria Ralstonia solanacearum en las etapas de germinación y plántula
La Calera, vol. 26, núm. 46, pp. 35-42, 2026
Universidad Nacional Agraria

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Recepción: 23 Febrero 2026
Aprobación: 20 Mayo 2026
Resumen: Las plantas silvestres de tomate (Solanum lycopersicum L.) constituyen un enorme reservorio de variabilidad genética, por lo que su utilización ha permitido avances significativos en el mejoramiento genético del tomate cultivado. El objetivo de esta investigación fue identificar poblaciones de tomate silvestres tolerantes a la bacteria Ralstonia solanacearum en las etapas de germinación y plántula, estableciéndolas como material base para programas de mejoramiento genético. Se inocularon 10 poblaciones de tomate con la cepa de Ralstonia solanacearum con código RsTB1. La prueba de germinación se realizó en una cámara de germinación ajustada a una temperatura constante de 27 ± 2 °C, con una humedad relativa del 70 ± 5 %. Las poblaciones se mantuvieron en oscuridad durante los primeros tres días y en condiciones ambientales del laboratorio durante siete días. El análisis de agrupamiento identificó seis poblaciones con tolerancia al patógeno (p ≤ 0.05). En ambas etapas fenológicas se identificaron poblaciones tolerantes, lo que sugiere que los mecanismos de tolerancia actúan de manera similar en cada etapa. Estos resultados destacan el potencial de poblaciones silvestres de tomate como material base para programas de mejoramiento genético orientados al manejo de la marchitez bacteriana.
Palabras clave: tomate silvestre, tolerancia, mejoramiento genético, resistencia, índice de germinación, radícula.
Abstract: Wild tomato plants (Solanum lycopersicum L.) constitute an enormous reservoir of genetic variability, and their use has led to significant advances in the genetic improvement of cultivated tomatoes. The objective of this research was to identify wild tomato populations tolerant to Ralstonia solanacearum in the germination and seedling stages, establishing them as base material for genetic improvement programs. Ten tomato populations were inoculated with the Ralstonia solanacearum strain coded RsTB1. The germination test was performed in a germination chamber set at a constant temperature of 27 ± 2 °C with a relative humidity of 70 ± 5%. The populations were kept in darkness for the first three days and under laboratory conditions for seven days. Cluster analysis identified six populations with tolerance to the pathogen (p ≤ 0.05). Tolerant populations were identified in both phenological stages, suggesting that tolerance mechanisms act similarly at each stage. These results highlight the potential of wild tomato populations as source material for genetic improvement programs aimed at managing bacterial wilt.
Keywords: Wild tomato, tolerance, genetic improvement, resistance, germination rate, radicle.
El tomate (Solanum lycopersicum L.) es la segunda hortaliza más cultivada y de mayor importancia alimenticia a nivel mundial por su contribución a la seguridad alimentaria y su amplio uso en la dieta humana (Kim et al., 2016). En el 2023, la producción global superó los 192.32 millones de toneladas cosechadas en una superficie de 5.4 millones de hectáreas (Organización de las Naciones Unidad para la Alimentación y la Agricultura [FAO], 2023). Debido a su amplia distribución mundial, este vegetal está frecuentemente expuesto tanto a factores bióticos como abióticos, los que disminuyen su rendimiento (Ramírez-Ojeda et al., 2021).
Entre los factores bióticos, la marchitez bacteriana causada por Ralstonia solanacearum representa una de las enfermedades más destructivas en solanáceas, particularmente en regiones tropicales y subtropicales (Valdez, 2016). Este patógeno presenta un amplio rango de hospederos, superior a 200 especies y una elevada variabilidad genética, lo que dificulta su manejo y favorece su persistencia en el suelo y en el agua (Khan et al., 2020; Yuliar et al., 2015). En el cultivo de tomate, las pérdidas asociadas a esta enfermedad pueden oscilar entre 0 y 90 %, dependiendo de las condiciones ambientales y del manejo agronómico (Yuliar et al., 2015).
La complejidad del patosistema (parasitismo de un subsistema ecológico) R. solanacearum-tomate se ve reforzada por la capacidad del patógeno para colonizar tejidos vasculares, persistir de forma endofítica y diseminarse a través de material vegetal, agua y suelo (Obrador-Sánchez et al., 2017; Jose et al., 2023). Estas características, sumadas a la limitada eficacia de los métodos de control químico y cultural, hacen que el desarrollo de materiales con tolerancia genética sea una de las estrategias más sostenibles para el manejo de la enfermedad (Karim y Hossain, 2018).
Las poblaciones silvestres de tomate adquieren especial relevancia, ya que constituyen un importante reservorio de variabilidad genética. Estudios sobre la historia de domesticación indican que el tomate cultivado se originó a partir de Solanum pimpinellifolium L. y Solanumlycopersicum var. cerasiforme, procesos que implicaron una reducción progresiva de la diversidad genética, incrementando la susceptibilidad a enfermedades en los materiales modernos (Razifard et al., 2020; Darwin et al., 2003). Por ello, los parientes silvestres representan una fuente de genes asociados a tolerancia frente a patógenos.
La evaluación de la tolerancia a R. solanacearum en etapas tempranas del desarrollo, como la germinación y la fase de plántula, es una alternativa para identificar materiales promisorios en un periodo corto de tiempo y bajo condiciones controladas (Deanda-Tovar et al., 2025; Kim et al., 2016). Estas etapas permiten detectar respuestas fisiológicas y de crecimiento asociadas a la interacción hospedante-patógeno, facilitando la selección preliminar de genotipos con mejor desempeño sanitario. El objetivo de este estudio fue identificar poblaciones silvestres de tomate con tolerancia a Ralstoniasolanacearum durante las etapas de germinación y plántula, que permita continuar con un programa de mejoramiento genético orientados al manejo de la marchitez bacteriana en este cultivo.
MATERIALES Y MÉTODOS
Ubicación del estudio
El estudio se efectuó en el laboratorio de la microbiología, de la Universidad Nacional Agraria localizada en las coordenadas 12°08’52’’ de latitud Norte y 86°09’41’’ de longitud Oeste, ubicada en el kilómetro 12.5 Carretera Norte, Managua, Nicaragua.
Material vegetal
Se evaluaron 10 poblaciones de tomate silvestres obtenidas del banco de germoplasma del Instituto Nicaragüense de Tecnología Agropecuaria (INTA); colectadas en diferentes departamentos de Nicaragua y la variedad INTA-JL5, denominada en adelante como JL5.
| Poblaciones | Lugar de colecta | Departamento | Municipio | Latitud Norte | Longitud Oeste |
| CJi | La Concordia | Jinotega | La Concordia | 13°11´45" | 86°10´00" |
| SEst1 | Sabana 1 | Estelí | Estelí | 13°08´51" | 86°36´30" |
| RR | Centro de Desarrollo Tecnológico El Recreo | RACCS | El Rama | 12°09´35" | 84°13´11" |
| JNS | Jicaro | Nueva Segovia | Jicaro | 13°40´15" | 86°09´31" |
| MM | Matiguás | Matagalpa | Matiguás | 12°50´14" | 85°27´43" |
| SRJ | San Rafael | Jinotega | San Rafael | 13°13´55" | 86°04´38" |
| SEst2 | Sabana 2 | Estelí | Estelí | 13°08´51" | 86°36´30" |
| SCh | Reserva de recursos genéticos Apacunca | Chinandega | Somotillo | 13°02´40" | 86°54´18" |
| TCh | Reserva de recursos genéticos Apacunca | Chinandega | Tomotillo | 13°02´40" | 86°54´18" |
| INTA | Centro Nacional de Investigación Agropecuaria | Managua | Managua | 12°8´21.43" | 86°6´47.44" |
| JL5 | Centro Nacional de Investigación Agropecuaria | Managua | Managua | 12°8´21.43" | 86°16´47.44" |
Se utilizó la cepa R. solanacearum (RsTb1) proporcionada por el laboratorio de microbiología de la Universidad Nacional Agraria (Nicaragua). El aislado bacteriano se cultivó en placas de agar nutritivo por un periodo de tres días a temperatura de 28 °C. Las unidades formadoras de colonias (UFC), se separaron del medio de cultivo agregando 10 ml de agua destilada estéril, raspando suavemente la superficie con un asa bacteriológica de plástico estéril. La suspensión se filtró a través de una gasa estéril en un vaso de precipitado previamente esterilizado. Las concentraciones celulares, se ajustaron a UFC mediante escala McFarland a una concentración de 1x108 UFC ml-1 (Gattoni et al., 2023; Kim et al., 2016).
La cepa se reactivó en agar nutritivo (AN) a 25 °C durante 24 horas. Se preparó una suspensión bacteriana siguiendo el método descrito por Kim et al. (2016).
Desinfección de las semillas
Las semillas fueron sumergidas en una placa de Petri con alcohol histológico al 95 % durante tres minutos, seguido de dos lavados con agua destilada estéril para eliminar el alcohol de las semillas. Luego fueron colocadas sobre un papel toalla estéril en una cámara de flujo laminar y se dejaron secar durante una hora, según la metodología del International Seed Testing Association (ISTA, 2026).
Diseño experimental
El experimento se realizó de octubre a noviembre del 2025 en una cámara de germinación (Lab-Tech Inc®, modelo D-7140, Hopkinton, MA, EE. UU.), en condiciones de oscuridad a 72 ± 2 °C y 70 ± 5 % de humedad relativa durante los tres primeros días; y en condiciones de luz y temperatura ambiental durante siete días. Para cada población se estableció una unidad experimental compuesta por 20 semillas, dispuestas sobre papel filtro en placas de Petri de 5 cm de diámetro. A cada unidad se añadió 2 ml de suspensión del inóculo de R. solanacearum, preparada previamente en condiciones estériles. Como testigo, se utilizaron unidades experimentales idénticas en las que se aplicaron 2 ml de agua destilada en lugar del inóculo, con el objetivo de determinar un efecto directo de la bacteria en la germinación. Posteriormente, cada placa se humedeció con 1 ml de agua destilada en tres momentos (3, 6 y 9 días después de la siembra), para mantener la humedad necesaria durante el ensayo. Se utilizó un Diseño Completamente al Azar con tres repeticiones.
Variables evaluadas
Índice de germinación e índice de daño. Se determinaron según las ecuaciones propuestas por Maguire (1962), quien señala que el índice de germinación (GRI) es igual a:
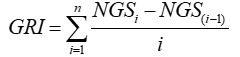
Donde i = 1, 2 . . . n; n = 10, número de recuentos realizados; NGSi, NGS (i-1) = número de semillas germinadas en los recuentos i (tiempo inicial), e i – 1 (tiempo final).
Y que el índice de daño (DRI) es iguala a:
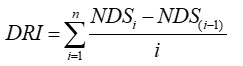
Donde i = 1, 2 . . . n; n = 10, número de recuentos realizados; NDSi, NDS (i-1) = número de semillas dañadas en los recuentos i (tiempo inicial), e i – 1 (tiempo final). El daño en la semilla se refiere a manchas y/o textura blanda.
Incidencia de Ralstonia en semilla (IR). Se determinó considerando la relación del entre semillas dañadas y total de semilla expresada en porcentaje como lo indica Maguire (1962).

A los 10 días después de la inoculación (DDI), se evaluaron variables de plántula como longitud de la radícula (cm), longitud de tallo (cm), longitud total de plántula (cm), peso húmedo (g) y peso seco (g) de plántula, para ello se obtuvo una muestra de cinco plántulas por unidad experimental, para las primeras tres variables se empleó una regla milimétrica; para determinar el peso húmedo y peso seco, se utilizó una balanza analítica (OHAUS PX323/E (320 g ± 0,001 mg; OHAUS Corp., Parsippany, NJ, EE. UU.), y para el secado de las muestras, un horno (J.P.Selecta, s.a), modelo Dry‑Big 288 serie 0541225, España).
Análisis de datos. Se realizó un análisis de conglomerados utilizando las distancias de Gower, (1967) y el algoritmo de Varianza Mínima de Ward, (1963), con el índice de germinación, índice de daños, incidencia, longitud de radícula, longitud de tallo, longitud total, peso húmedo y peso seco. Se utilizó una altura de corte de conglomerado de 2.2 (Hair et al., 2019). Las variables fueron sometidos a pruebas de homogeneidad de varianza y normalidad, se aplicó un Análisis de Varianza y comparaciones de medias de Fisher-LSD (Diferencia Mínima Significativa), a los índices de germinación y de daño, seleccionando el modelo de mejor ajuste, es decir con el menor valor o Índice de Akaike (AIC), considerando los grupos y las poblaciones anidadas dentro de los grupos como fuentes de varianza. El análisis se realizó utilizando el software estadístico R, versión 4.5.1 (R Core Team, 2025).
RESULTADOS Y DISCUSIÓN
Respuesta a Ralstonia solanacearum durante la germinación
El análisis de agrupamiento jerárquico permitió organizar las poblaciones en cuatro grupos con comportamiento contrastante durante la germinación bajo inoculación con R. solanacearum. La separación entre grupos estuvo determinada por diferencias en germinación, semillas dañadas e incidencia de la enfermedad. El análisis de agrupamiento identificó al segundo grupo con el mayor porcentaje de poblaciones (61 %), con los mayores valores para semillas germinadas y baja incidencia del patógeno, mientras que los restantes incluyeron menor número de poblaciones con mayor afectación (Figura 1).
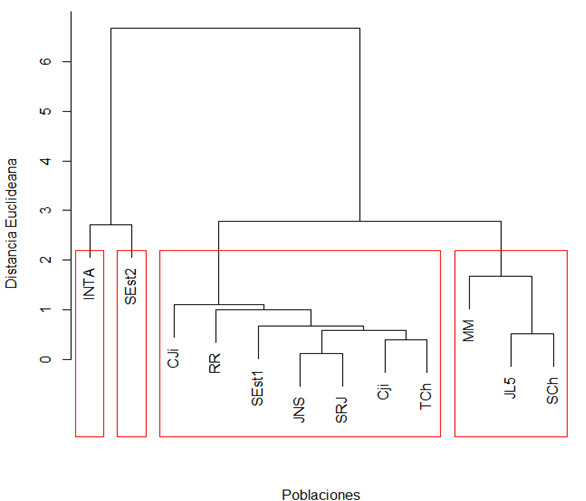
Figura 1.
Dendrograma jerárquico generado utilizando distancias de Gower y el algoritmo de Varianza Mínima de Ward basado en índices de germinación, índice de daño e incidencia para 10 poblaciones de tomate silvestre inoculados con Ralstonia solanacearum en fase de germinación.
El análisis de componentes principales (ACP) verificó la clasificación obtenida por el análisis de conglomerados y explica una varianza del 72.7 % y del 27.3 %, respectivamente; juntas, explicaron el 100 % de la varianza en los datos (Figura 2). Este análisis también muestra que las poblaciones del grupo tres presentaron alta incidencia y un alto índice de semillas dañadas, mientras que las poblaciones del grupo cuatro no tienen relación con la variable índice de semillas germinadas. El componente principal uno aisló la mayor incidencia e índice de semillas dañadas, sin embargo, el componente principal dos agrupa el índice de semillas germinadas, como variable explicativa.
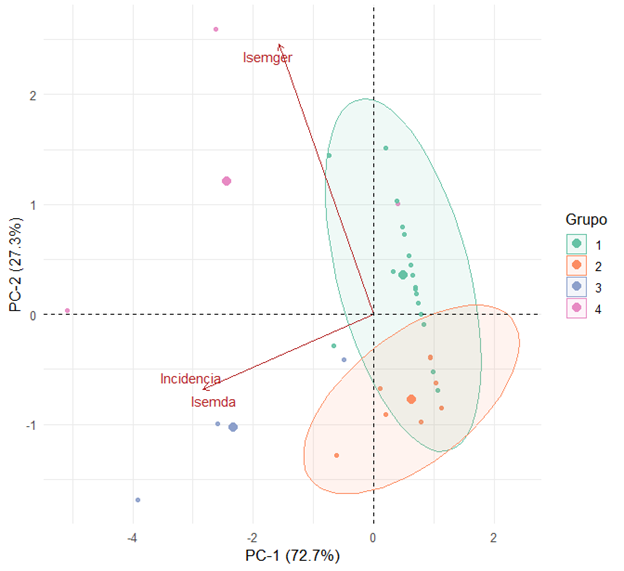
Figura 2.
Análisis de componentes principales para10 poblaciones de tomate silvestre inoculados con Ralstonia solanacearum en fase de germinación. Isemda: índice de semillas dañadas, Isemger: índice de semillas germinadas.
La población definida como INTA registró el mayor número de semillas germinadas (μ=11.12), lo que muestra mayor capacidad para sostener el proceso germinativo bajo presión del patógeno. Las poblaciones Cji, RR, SEst1, JNS, SRJ y TCh presentaron 0 % de incidencia y menor número de semillas dañadas, combinando germinación intermedia con baja afectación por el patógeno. Este comportamiento los ubica entre los materiales con mejor desempeño en esta etapa. Por el contrario, las poblaciones JL5, MM y SEst2 concentraron los valores más altos de incidencia (11.33 %, 10 % y 9.26 %, respectivamente y mayor número de semillas dañadas (Cuadro 2). En estas poblaciones, la disminución en la germinación coincidió con un incremento en el daño observado, lo que sugiere mayor impacto del patógeno durante el desarrollo inicial. Los coeficientes (AIC), reportados constituyen al mejor ajuste en el modelo empleado para explicar el comportamiento de las poblaciones sometidas a presión por la bacteria (Ralstonia solanacearum). En este caso, la variable índice de semilla dañada se constituye en un indicador de importancia para la identificación de poblaciones promisorias.
En algunas poblaciones se observaron semillas dañadas con incidencia baja o nula, lo que puede asociarse a infección superficial sin comprometer completamente la viabilidad del embrión, situación descrita en otras solanáceas (Sharma y Singh, 2015). El comportamiento registrado se podría justificar considerando reportes que señalan que R. solanacearum interfiere en la germinación mediante la acción de enzimas hidrolíticas que afectan la integridad celular y la absorción de agua (Baroukh et al., 2022; Murti et al., 2021). Sin embargo, la magnitud del efecto varió entre poblaciones, lo que confirma que la respuesta depende del genotipo como lo indican Bhunchoth et al. (2025).
Los resultados en esta investigación permiten deducir que, la fase de germinación es determinante para la expresión de tolerancia o susceptibilidad a R. solanacearum y permite identificar poblaciones con potencial para estudios posteriores orientados a la caracterización de mecanismos de tolerancia o resistencia en diferentes fases de su ciclo biológico.
| Poblaciones | Índice de semillas germinadas | Índice de semillas dañadas | Incidencia (%) |
| INTA | 11.12 a | 2.72 bcd | 3.20 bcd |
| RR | 6.61 ab | 0.96 a | 0.00 a |
| JNS | 5.15 ab | 1.56 a | 0.00 a |
| SRJ | 4.83 ab | 1.62 ab | 0.00 a |
| CJi | 4.36 ab | 1.85 bc | 1.63 bc |
| TCh | 3.85 bc | 1.88 bc | 0.83 a |
| SEst2 | 3.78 bc | 1.63 bcd | 9.26 b |
| SEst1 | 3.28 bcd | 1.96 bcd | 0.00 a |
| MM | 2.21 bcd | 2.92 cde | 10.00 cde |
| SCh | 1.43 cdf | 1.30 a | 0.00 a |
| JL5 | 0.00 b-e | 3.81 c-f | 11.33 cde |
| P (0.05) | 0.0001 | 0.0006 | 0.0003 |
| CV (%) | 40.07 | 105.29 | 127.86 |
| AIC | 274.93 | 25.45 | 320.08 |
El análisis detectó diferencias (p ≤ 0.05) en las fuentes de variación para todos los índices evaluados, registrando coeficientes de variación con valores altos (> 40 %), sim embargo, por tratarse de un experimento en el que las poblaciones fueron sometidas a un alto estrés, Paredes-Cervantes et al. (2025), indican que esta condición aumenta la variación aleatoria.
Respuesta a Ralstonia solanacearumdurante la fase de plántula
Una forma rápida de evaluar la resistencia de las poblaciones a los microorganismos causantes de enfermedades es exponer las plántulas a concentraciones de inóculo (Arunakumar y Gnanesh, 2023). El análisis de agrupamiento identificó las poblaciones en cuatro grupos con respuestas contrastantes durante la fase de plántula (Figura 3). La divergencia entre los grupos estuvo determinada por diferencias en longitud de radícula (cm), longitud de tallo (cm), peso húmedo de plántula (g) y peso seco de plántula (g). Los grupos jerárquicos 2 y 4 concentraron el mayor porcentaje de poblaciones (80 %) con los mayores valores para longitud de radícula, longitud de tallo, peso húmedo, peso seco y peso total de plántula.
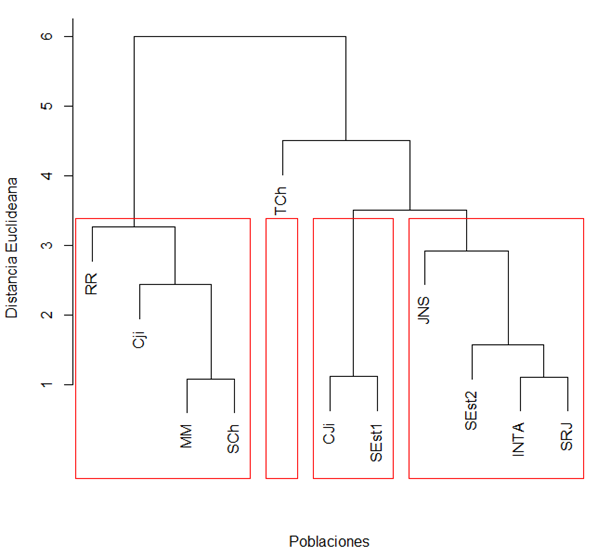
Figura 3.
Dendrograma jerárquico generado utilizando distancias de Gower y el algoritmo de Varianza Mínima de Ward basado en longitudes de radícula, longitud de tallo y biomasa para 10 poblaciones de tomate silvestre inoculados con Ralstonia solanacearum en fase de plántula.
El análisis de componentes principales (ACP) verificó la clasificación obtenida por el análisis de conglomerados y explica una varianza del 51 % y del 32 %, respectivamente; juntas explicaron el 83 % de la varianza total (Figura 4). El ACP muestra que las poblaciones uno y dos están asociadas positivamente a las variables longitud de radícula, longitud de tallo, longitud total, esto indica que las plántulas acumulan mayor cantidad de biomasa y las poblaciones del grupo dos están asociadas positivamente al peso húmedo y peso seco con mayor tejido denso. Estos parámetros suelen relacionarse con plantas de alto vigor fisiológico y aptas para el establecimiento en campo.
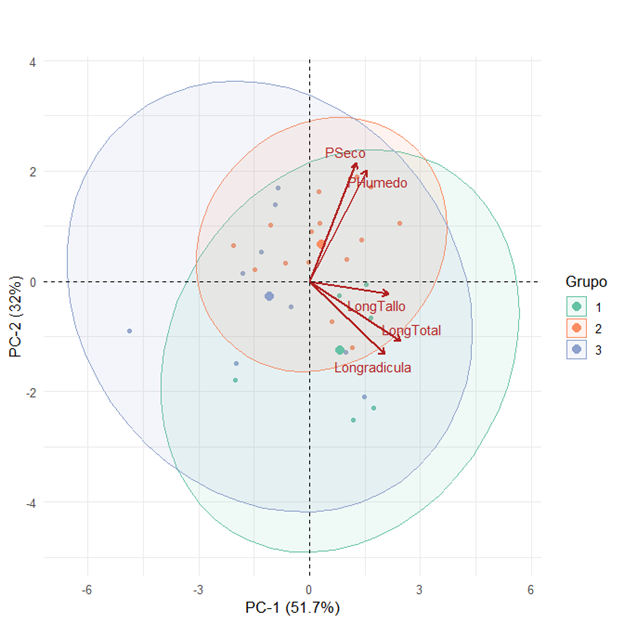
Figura 4.
Análisis de componentes principales para10 poblaciones de tomate inoculados en fase de plántula con Ralstonia solanacearum.
Las poblaciones, INTA, JNS, MM y TCH presentan los mejores valores para longitud de radícula, longitud de tallo y longitud total, comportamiento que no se correlaciona con el peso húmedo y seco (Cuadro 3). Pérez y Villareal, (2024), registraron incrementos superiores en la longitud del tallo, acompañados de aumentos en los pesos fresco y seco del tejido aéreo en plántulas de tomate; lo que demuestra que el comportamiento de las variables de crecimiento puede variar según el fenotipo, la fase fenológica del cultivo y las condiciones experimentales como lo plantea Machado et al. (2024).
Las poblaciones con los menores valores de longitud de radícula, longitud de tallo y longitud total mantienen, en la mayoría de las poblaciones, un peso seco igual al de las poblaciones con los mejores valores de longitud de radícula, longitud de tallo y longitud total.
Las raíces más pequeñas tienen menor capacidad de absorción de agua y nutrientes, lo que se refleja en un crecimiento general reducido, sin embargo, en este estudio, no se traduce en menor peso húmedo y seco, lo que podría atribuirse a un diámetro de tallo mayor. La reducción en longitud radicular y peso en las poblaciones más afectadas implica menor capacidad de exploración del sustrato y absorción de agua y nutrientes, lo que puede limitar el establecimiento posterior del cultivo. Hossain et al. (2021), describen reducciones significativas en el crecimiento radicular y aéreo tras la inoculación con R. solanacearum.
En este estudio, se observa en algunas poblaciones mayor susceptibilidad a la bacteria durante la germinación que en la etapa de plántula. Esto puede deberse a que, en la germinación, la semilla está iniciando su actividad metabólica y depende totalmente de la absorción de agua para emerger, como lo indica Upretee et al. (2024). En esta fase (germinación), cualquier alteración causada por el patógeno puede afectar directamente el proceso de emergencia; en cambio, en la etapa de plántula los tejidos ya están formados y la planta tiene mayor desarrollo estructural, lo que puede reducir la manifestación visible del daño.
| Poblaciones | Longitud de radícula (cm) | Longitud de tallo (cm) | Longitud Total (cm) | Peso húmedo (g) | Peso seco (g) |
| INTA | 3.19 a | 2.72 abc | 5.90 a | 0.24 b-e | 0.023 ab |
| JNS | 2.57 a | 1.56 efg | 4.16 b-e | 0.61 a | 0.020 abc |
| MM | 2.27 ab | 2.92 a | 5.16 ab | 0.15 b-e | 0.010 cd |
| TCH | 2.21 a-d | 1.88 def | 4.10 b-f | 0.10 cde | 0.003 ef |
| SEst2 | 1.82 b-f | 1.63 efg | 3.47 c-g | 0.35 b | 0.026 a |
| SRJ | 1.44 c-g | 1.62 efg | 3.06 c-g | 0.25 b-e | 0.023 ab |
| SCh | 1.44 c-g | 1.30 fg | 2.73 d-g | 0.10 cde | 0.013 cd |
| SEst1 | 1.08 efg | 1.96 c-f | 3.05 c-g | 0.18 b-e | 0.020 abc |
| RR | 0.97 fg | 0.96 g | 1.90 g | 0.26 bcd | 0.020 ab |
| CJi | 0.60 g | 1.85 def | 2.46 fg | 0.21 b-e | 0.020 abc |
| p (0.05) | 0.0056 | 0.0006 | 0.0015 | 0.0023 | 0.0003 |
| CV (%) | 37.64 | 25.45 | 27.20 | 57.68 | 31.29 |
| AIC | 139.65 | 105.29 | 190.31 | 56.07 | 443.98 |
CONCLUSIONES
Existe variabilidad en la tolerancia a Ralstonia solanacearum entre las poblaciones; las mejores corresponden a INTA, RR, JNS y SRJ, CJi y TCh; con desempeño superior durante la germinación y fase de plántula, las que se constituyen en material genético promisorio para programas de mejoramiento.
REFERENCIAS BIBLIOGRÁFICAS
Arunakumar, G. S., & Gnanesh, B. N. (2023). Evaluation of artificial inoculation methods to determine resistance reaction to dry root rot and black root rot disease in mulberry (Morus spp.). Archives of Phytopathology and Plant Protection, 56(1), 49–65. https://doi.org/10.1080/03235408.2023.2170692
Baroukh, C., Zemouri, M., & Genin, S. (2022). Trophic preferences of the pathogen Ralstonia solanacearum and consequences on its growth in xylem sap. MicrobiologyOpen, 11(1), e1240. https://doi.org/10.1002/mbo3.1240
Bhunchoth, A., Poncheewin, W., Yongsuwan, A., Chiangta, J., Thunnom, B., Aesomnuk, W., & Ruanjaichon, V. (2025). Genome-wide association analysis and breeding-oriented SNP marker development for bacterial wilt resistance in tomato (Solanum lycopersicum L.). Plants, 14(19), 3036. https://doi.org/10.3390/plants14193036
Deanda-Tovar, A. A., Rodríguez-Pérez, J. E., Sahagún-Castellanos, J., Colinas-y-León, M. T. B., Pérez-Rodríguez, P., & Paredes Cervantes, A. E. (2025). Tomato lines tolerant to sodium chloride at early growth stages. Horticulturae, 11, 532. https://doi.org/10.3390/horticulturae11030532
Darwin, S. C., Knapp, S., & Peralta, L. E. (2003). Taxonomy of tomatoes in the Galápagos Islands: Native and introduced species of section Solanum Lycopersicon (Solanaceae). Systematics and Biodiversity, 1(1), 29–55. https://doi.org/10.1017/S1477200003001019
Food and Agriculture Organization of the United Nations. (2023). FAOSTAT statistical database. https://www.fao.org/faostat
Gower, J. C. (1967). A comparison of some methods of cluster analysis. Biometrics, 23(2), 623–637. https://doi.org/10.2307/2528417
Gattoni, K. M., Park, S. W., & Lawrence, K. S. (2023). Evaluation of the mechanism of action of Bacillus spp. to manage Meloidogyne incognita with split root assay RT-qPCR and qPCR. Frontiers in Plant Science, 13, 1-13. https://doi.org/10.3389/fpls.2022.1079109
Hossain, M. F., Billah, M., Ali, M. R., Parvez, M. S. A., Zaoti, Z. F., Hasan, S. Z., & Sikdar, B. (2021). Molecular identification and biological control of Ralstonia solanacearum from wilt of papaya by natural compounds and Bacillus subtilis: an integrated experimental and computational study. Saudi Journal of Biological Sciences, 28(12), 6972-6986. https://pmc.ncbi.nlm.nih.gov/articles/PMC8626333/
Hair, J. F., Black, W. C., Babin, B. J., & Anderson, R. E. (2019). Multivariate data analysis (8.ª ed.). Cengage Learning. https://eli.johogo.com/Class/CCU/SEM/_Multivariate%20Data%20Analysis_Hair.pdf
International Seed Testing Association. (2026). International Rules for Seed Testing. ISTA. https://www.seedtest.org/api/rm/REC7F8M33NAQ269/introduction-to-the-ista-rules-2.pdf
Jose, J., Éva, C., Bozsó, Z., Hamow, K. Á., Fekete, Z., Fabián, A., & Sági, L. (2023). Global transcriptome and targeted metabolite analyses of roots reveal different defence mechanisms against Ralstonia solanacearum infection in two resistant potato cultivars. Frontiers in Plant Science, 13, 1-19. https://doi.org/10.3389/fpls.2022.1065419
Kim, S. G., Hur, O. S., Ro, N. Y., Ko, H. C., Rhee, J. H., Sung, J. S., Ryu, K. Y., Lee, S. Y., & Baek, H. J. (2016). Evaluation of resistance to Ralstonia solanacearum in tomato genetic resources at seedling stage. Plant Pathology Journal, 32(1), 58–64. https://doi.org/10.5423/PPJ.NT.06.2015.0121
Khan, R. A. A., Najeeb, S., Mao, Z., Ling, J., Yang, Y., Li, Y., & Xie, B. (2020). Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic bacteria and root-knot nematode. Microorganisms, 8(3), 401. https://doi.org/10.3390/microorganisms8030401
Karim, Z., & Hossain, M. (2018). Management of bacterial wilt (Ralstonia solanacearum) of potato: focus on natural bioactive compounds. Journal of Biodiversity Conservation and Bioresource Management, 4(1), 73–92. https://doi.org/10.3329/jbcbm.v4i1.37879
Murti, R. H., Afifah, E. N., & Nuringtyas, T. R. (2021). Metabolomic response of tomatoes (Solanum lycopersicum L.) against bacterial wilt (Ralstonia solanacearum) using 1H-NMR spectroscopy. Plants, 10(6), 1143. https://doi.org/10.3390/plants10061143
Machado, J., Heuvelink, E., Vasconcelos, M. W., Cunha, L. M., Finkers, R., & Carvalho, S. M. P. (2024). Exploring tomato phenotypic variability under combined nitrogen and water deficit. Plant and Soil, 496(1-2), 123–138. https://doi.org/10.1007/s11104-023-06023-5
Maguire, J. D. (1962). Speed of germination Aid in selection and evaluation for seedling emergence and vigor. Crop Science, 2(2), 176–177. https://doi.org/10.2135/cropsci1962.0011183X000200020033X
Obrador-Sánchez, J. A., Tzec-Simá, M., Canto-Canché, B., & Higuera-Ciapara, I. (2017). Técnicas para el aislamiento, identificación y caracterización molecular de cepas de Ralstonia solanacearum relacionadas a moko del plátano. Revista Mexicana de Fitopatología, 35(3), 509–533. https://doi.org/10.18781/r.mex.fit.1705-1
Paredes-Cervantes, A. E., Rodríguez-Pérez, J. E., Sahagún-Castellanos, J., Leyva-Mir, S. G., Hernández-Rodríguez, M., & Deanda-Tovar, A. A. (2025). Response of Solanum lycopersicum L. to Fusarium oxysporum during germination and seedling stages. Agronomy, 15(9), 1-17. https://doi.org/10.3390/agronomy15092089
Pérez, D., Ramírez-Rodríguez, H., & Villarreal-Quintanilla, J. A. (2024). Bioestimulantes de plantas del semidesierto en el crecimiento radicular y aéreo de plántulas de tomate. Agropecuaria, 11(3), e4145. https://doi.org/10.19136/era.a11n3.4145
Ramírez-Ojeda, G., Peralta, I. E., Rodríguez-Guzmán, E., Chávez-Servia, J. L., Sahagún-Castellanos, J., & Rodríguez-Pérez, J. E. (2021). Climatic diversity and ecological descriptors of wild tomato species (Solanum sect. Lycopersicon) and close related species (Solanum sect. Juglandifolia y sect. Lycopersicoides) in Latin America. Plants, 10(5), 1-17. https://doi.org/10.3390/plants10050855
Razifard, H., Ramos, A., Della Valle, A. L., Bodary, C., Goetz, E., Manser, E. J., Li, X., Zhang, L., Visa, S., Tieman, D., van der Knaap, E., & Caicedo, A. L. (2020). Genomic evidence for complex domestication history of the cultivated tomato in Latin America. Molecular Biology and Evolution, 37(4), 1118–1132. https://doi.org/10.1093/molbev/msz297
R Core Team. (2025). R: A language and environment for statistical computing (Version 2025) [Software]. R Foundation for Statistical Computing. https://www.R-project.org/
Sharma, D. K., Mehta, S., & Singh, R. (2015). Incidence and colonisation of Ralstonia solanacearum (Smith) Yabuuchi et al. in brinjal (Solanum melongena L.) seeds in Rajasthan, India. Journal of Phytological Research, 28(1 & 2), 71–79. https://www.jphytolres.org/system/files?file=old_papers%2F10.Dilip+Kumar+Sharma%C2%AC2+%282%29.pdf
Upretee, P., Bandara, M. S., & Tanino, K. K. (2024). The role of seed characteristics on water uptake preceding germination. Seeds, 3(4), 559–574. https://doi.org/10.3390/seeds3040038
Valdez, M. M. (2016). Bacterias antagonistas para el control biológico de Ralstonia solanacearum (E. F. Smith) en tomate (Solanum lycopersicum L.) [Tesis de maestría, Centro de Investigación en Alimentación y Desarrollo]. Repositorio Institucional. https://ciad.repositorioinstitucional.mx/jspui/bitstream/1006/776/1/Valdez-Morales%20M%20T_MC_2016.pdf
Ward, J. H. (1963). Hierarchical grouping to optimize an objective function. Journal of the American Statistical Association, 58(301), 236–244. https://doi.org/10.1080/01621459.1963.10500845
Yuliar Nion, Y. A., & Toyota, K. (2015). Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes and Environments, 30(1), 1–11. https://doi.org/10.1264/jsme2.ME14144
Notas de autor
Información adicional
redalyc-journal-id: 7858