Recursos Naturales y Ambiente
Consorcio de hongos entomopatógenos para el manejo de termitas Blattodea: Isóptera (Nasutitermes corniger)
Consortium of entomopathogenic fungi for the management of Blattodea: Isoptera termites (Nasutitermes corniger)


Consorcio de hongos entomopatógenos para el manejo de termitas Blattodea: Isóptera (Nasutitermes corniger)
La Calera, vol. 26, núm. 46, pp. 43-51, 2026
Universidad Nacional Agraria

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Recepción: 11 Marzo 2026
Aprobación: 03 Junio 2026
Resumen: Las termitas son insectos polifórmicos, eusociales, exopterigotas que viven en termiteros, su importancia radica en los daños que pueden ocasionar en postes, árboles, cultivos, casas, obras de arte y muebles, con impacto global. El objetivo de este estudio fue evaluar un consorcio de entomopatógenos de los géneros Beauveria, Metarhizium, Lecanicillium y Paecilomyces en la concentración de 1x108 esporas ml-1. La investigación se desarrolló en el Campus Juanico, Universidad de Oriente, Monagas. Se evaluó un testigo ─solo agua─ y cuatro dosis del consorcio con dosis de 5 ml L-1, 10 ml L-1, 20 ml L-1 y 30 ml L-1. Se utilizó un diseño completamente aleatorizado con cinco repeticiones y la unidad experimental constituida por el termitero. Se cuantificó, el número de termitas vivas y muertas por termitero, termitas muertas colonizadas, porcentaje de colonización y peso de 100 termitas expresadas en gramos. Los datos se analizaron por estadística: descriptiva y no paramétrica (Kruskal Wallis), y comparados por suma de rangos. Se observó ajuste lineal de regresión, entre el volumen y el peso de las termitas; diferencias entre tratamientos, para el número de termitas vivas y termitas muertas, con menos termitas vivas en las concentraciones de 20 ml L-1 y 30 ml L-1 a los 25 después de la aplicación y a nivel de campo y cero termitas vivas, en todas las concentraciones, a los 50 días después de la aplicación; cero termitas muertas a los 25 días, y a los 50 días, mayor número de termitas muertas en las concentraciones de 20 y 30. El tratamiento testigo se mantuvo estable ante la aplicación del biológico. Las mayores diferencias en el número de termitas muertas colonizadas y el porcentaje de colonización se observaron a favor de las concentraciones de 20 ml L-1 y 30 ml L-1. El peso de 100 termitas no varió entre los distintos tratamientos. La formulación de los entomopatógenos constituye una tecnología potencial para el manejo del insecto plaga en condiciones naturales.
Palabras clave: Insecticida biológico, Blattodea: Isoptera, control biológico, manejo sostenible de plagas.
Abstract: Termites are polymorphic, eusocial, exopterygote insects that live in termite mounds. Their importance lies in the damage they can cause to poles, trees, crops, houses, works of art, and furniture, with a global impact. The objective of this study was to evaluate a consortium of entomopathogens from the genera Beauveria, Metarhizium, Lecanicillium, and Paecilomyces at a concentration of 1 x 10⁸ spores ml⁻¹. The research was conducted at the Juanico Campus of the University of Oriente, Monagas. A control (water only) and four doses of the consortium were evaluated at 5 ml L⁻¹, 10 ml L⁻¹, 20 ml L⁻¹, and 30 ml L⁻¹. A completely randomized design with five replicates was used, and the experimental unit was the termite mound. The number of live and dead termites per mound, colonized dead termites, colonization percentage, and weight of 100 termites expressed in grams were quantified. The data were analyzed using descriptive and non-parametric statistics (Kruskal-Wallis test) and compared by rank-sum test. A linear regression fit was observed between termite volume and weight; differences between treatments were found for the number of live and dead termites, with fewer live termites at concentrations of 20 ml L-1 and 30 ml L-1 25 days after application and in the field, and zero live termites at all concentrations 50 days after application; zero dead termites at 25 days, and at 50 days, a higher number of dead termites at concentrations of 20 and 30 ml L-1. The control treatment remained stable after the application of the biological agent. The greatest differences in the number of colonized dead termites and the percentage of colonization were observed with the 20 ml L-1 and 30 ml L-1 concentrations. The weight of 100 termites did not vary among the different treatments. The formulation of entomopathogens represents a potential technology for managing this insect pest under natural conditions.
Keywords: Biological insecticide, Blattodea: Isoptera, biological control, sustainable pest management.
Las termitas (orden Blattodea: infraorden Isoptera) constituyen un grupo de insectos eusociales de distribución cosmopolita, con más de 3 000 especies descritas, agrupadas en diversas familias, ─Mastotermitidae, Archotermopsidae, Stolotermitidae, Kalotermitidae, Rhinotermitidae, Stylotermitidae, Hodotermitidae, Serritermitidae y Termitidae─ ; su importancia radica en los daños que pueden ocasionar a postes, árboles, cultivos, casas, obras de arte y muebles, con un impacto global estimado en 40 000 millones de dólares anuales (Rust y Su, 2012; Ricart et al., 2015).
Las termitas son excelentes descomponedores, se alimentan de restos de plantas, incluyendo madera (Mugerwa, 2015). Son capaces de atacar muchos cultivos agrícolas como leguminosas, hortalizas, raíces y tubérculos, frutales, entre otros (Qasim et al., 2015); además, de gran diversidad de especies de árboles. Li et al. (2010) señalan que entre 40 % y 60 % de los árboles en áreas montañosas del sur de China, mueren por ataque de termitas; y afectan infraestructuras de madera (Hassan et al., 2024).
En Venezuela, Rosales (2001) señala a las familias Kalotermitidae, Rhinotermitidae y Termitidae como las de mayor importancia económica, en ellas se citan 29 géneros y 48 especies. En la familia Kalotermitidae se incluye Cryptotermes brevis Walker, una de las especies de mayor importancia, cosmopolita; de Rhinotermitidae, son conocidos: Heterotermes convexionotatus Snyder, como la especie más común y a Coptotermes testaceus L.; y de la familia Termitidae, con mayor número de especies en el país, es mencionado el género Nasutitermes spp. con varias especies de amplia distribución. Por otro lado, en el país, los trabajos de investigación sobre control de termitas son inexistentes ─aunque si se realizan en otros países─; de manera similar, no se dispone de un método de control biológico, evaluado en condiciones experimentales que contribuya a minimizar el efecto de estos insectos plaga. En consecuencia, poca información está disponible en relación con el uso de hongos entomopatógenos para el manejo de termitas a nivel de campo.
Entre las alternativas de control para estos insectos plagas son citadas estrategias químicas y biológicas, estas últimas, realizadas por hongos entomopatógenos (Morris, 2000; Méndez y Equihua 2001). Pérez-Levastida (2014) menciona que el control químico ha sido ineficaz para controlar termitas, debido a la bioecología y organización de estos insectos; en consecuencia, esto ha implicado la búsqueda de alternativas de control que tengan menor impacto en la salud pública y que sean efectivas.
La estrategia de uso de control biológico con organismos benéficos como los hongos entomopatógenos, es una opción ecológica y económicamente viable para la regulación de diferentes especies de termitas plaga (Butt et al., 2001; Moino y Alves, 1998; Ferron, 1978). Tanada y Kaya (1993) relatan que los primeros microorganismos encontrados causando enfermedades en isópteros, fueron hongos entomopatógenos, debido a su notable capacidad de invasión y su rápido crecimiento macroscópico sobre la superficie de sus hospederos.
Las enfermedades fungosas en insectos son comunes y en ocasiones lo suficientemente severas para causar epizootias ─enfermedades infectocontagiosas─ capaces de eliminar casi por completo una población en un hábitat determinado (Herrera y Ulloa, 1990). Los hongos entomopatógenos poseen ciertas ventajas sobre otros agentes de control microbiano ─bacterias, protozoarios y virus─, por su competencia para atacar todas sus fases; además, infectan a los que normalmente no son susceptibles a bacterias y virus. Entre ellos son citados Metarhizium anisopliae Sun. y Beauveria bassiana Vuillemin (Ferron, 1978).
El control biológico provee herramientas adicionales para el manejo de plagas que permite romper los ciclos de resistencia de los insectos, así como solventar problemas de residuos de agroquímicos para la producción de alimentos orgánicos (Fravel, 2005). Existen más de 750 especies de hongos entomopatógenos diseminados en el ambiente que provocan infecciones fungosas a ciertos insectos, las cuales pueden regular hasta en 80 % sus poblaciones (Lobo et al., 2016). Dentro de los géneros más importantes están: Metarhizium, Beauveria, Aschersonia, Entomophthora,Zoophathora, Hirsutella, Fusarium y Verticillium (Acuña-Jiménez et al., 2015).
Beauveria bassiana es un hongo entomopatógeno (Abdu-Allah et al., 2015), su modo de acción es por contacto (Alcalá-Gómez et al., 2017), contaminando la microflora de la cutícula del insecto, cuando ocurre la germinación de la espora sobre el insecto plaga y la hifa se introduce al integumento ─tejido que forma la pared externa de la cutícula del cuerpo de los insectos─ e ingresar al hemocele, después propagarse mediante la hemolinfa por todo el insecto, causa su muerte por las toxinas ─toxemia─ que el mismo secreta (Petlamul y Prasertsan, 2012). B. bassiana presenta una gran diversidad genética y, por lo regular, se desarrolla en hábitats agrícolas (Bidochka et al., 1998; Rustiguel et al., 2018).
Beauveria bassiana y Metarhizium anisopliae se combinan debido a su modo de acción, ambos actúan por contacto. El primero lo ejerce por una estrategia tóxica mediante oosporeinas e invade al hospedero; mientras que, el segundo lo realiza por una estrategia de crecimiento, con la formación de apresorios e invasión del hospedero, lo que se traduce en un control biológico mayor de plagas (Rustiguel et al., 2018). Otros estudios se han enfocado en establecer cuál de estos dos hongos tiene mejor efecto fungicida (Barbosa et al., 2018). Ambos hongos entomopatógenos infectan positivamente a las termitas (Rath, 2000), al invadir a su hospedero a través del integumento y causar la muerte por agotamiento de sus metabolitos, debido a la destrucción de tejidos vitales o combinación de ambos métodos (Wang y Powell, 2004).
Los hongos que atacan las termitas en condiciones naturales y/o en otros ambientes, se les considera una alternativa importante para su control biológico. En estudios in vitro se ha demostrado que hongos como Aspergillus flavus,Entomophthora sp. (Espero, 1984), Beauveria bassiana (Zoberi y Grace, 1990) y Paecilomyces fumosoroseus (Chai, 1995) poseen niveles importantes de patogenicidad para termitas; además, se ha registrado el hongo Metarhizium anisopliae, atacando termitas como Reticulitermes flavipes (Zoberi 1995) y Coptotermes formosanus (Sajap y Kaur, 1990) en condiciones naturales.
Los hongos entomopatógenos, particularmente de los géneros Metarhizium, Beauveria, Lecanicillium e Isaria, tienen diversas aplicaciones en el manejo de plagas (Rath, 2000). Estas especies son relativamente fáciles de producir en masa y altamente efectivos. Estudios realizados por Khan et al. (1990) y Khan et al. (1991) en termitas de cultivos agroforestales ─Odontotermes brunneus, O. wallonensis y O. obesos─ tratadas con los hongos: B bassiana, Metarhizium anisopliae var. anisopliae, M. flavoviridae var. minos, Paecilomyces lilacinus y P.fumosoroseus, indicaron a B. bassiana como la más patogénica.
En función de los anteriores hallazgos experimentales, la presente investigación tiene por objetivo evaluar dosis de ECOINSET, un consorcio de hongos entomopatógenos constituidos por Beauveria bassiana,Metarhizium anisopliae,Lecanicillium lecanii y Paecilomyces sp. para el control de termitas o comején.
MATERIALES Y MÉTODOS
Ubicación y establecimiento del experimento
El experimento se realizó en dos fases, a nivel de campo y en laboratorio, ambas en el Centro de Investigación y Postgrado, Campus Juanico de la Universidad de Oriente, en Maturín, Monagas, Venezuela; ubicado en las coordenadas 9º44′13″ de latitud Norte y 63º09′28″ de longitud Oeste.
En las áreas adyacentes del campus se seleccionaron de manera aleatoria 25 árboles de diferentes especies con termiteros, procurando la mayor similitud en tamaño y forma de los nidos, y las evaluaciones en laboratorio se realizaron en la Clínica Universitaria de Diagnóstico Agropecuario (CUDA).
Tratamientos
Consorcio de entomopatógenos (bioinsumo). La formulación del bioinsumo presenta cuatro cepas de hongos entomopatógenos: Beauveria bassiana, Metarhizium anisopliae, Lecanicillium lecanii y Paecilomyces sp., en concentración de 1x108 esporas ml L-1. Se evaluaron cinco tratamientos, constituidos por un tratamiento testigo (agua), a los termiteros y cuatro dosis del bioinsumo ECOINSET, en concentraciones de 5 ml L-1, 10 ml L-1, 20 ml L-1 y 30 ml L-1.
Aplicación del bioinsumo y muestreo. En los termiteros de los árboles seleccionados se realizaron tres aberturas en la parte superior para una mejor distribución del producto, por donde se aplicó dos litros de la solución del bioinsumo, para cada una de sus concentraciones, con un rociador, luego, se taparon. La perforación y posterior sellado de los orificios en los termiteros, se realizó de acuerdo con la metodología de Pérez-Levastida (2014), usando papel absorbente humedecido.
Transcurridos 25 días después de la aplicación de los tratamientos ─fase de campo─, los termiteros se retiraron de los árboles, separándolos en cuatro porciones de similar tamaño; dos de ellas, fueron evaluadas inmediatamente (según variables) y las otras dos partes, colocadas en cajas plásticas, mantenidas con papel absorbente humedecido y colocado en el fondo de la caja plástica para su traslado al CUDA ─fase de laboratorio─, para ser evaluadas 25 días después de su traslado, lo que correspondió a 50 días después de la aplicación del consorcio de entomopatógenos.
Las evaluaciones, a los 25 y a los 50 días después de la aplicación del consorcio de entomopatógenos, se efectuó de la siguiente manera: las dos partes del termitero se colocaron en el interior de una bolsa plástica y fueron agitadas manualmente, con la finalidad de separar los insectos plagas del resto de las partes del termitero.
Debido a la dificultad de cuantificar directamente el número de individuos en colonias densas de Isóptera, se construyó un modelo de regresión lineal simple basado en la relación entre el volumen aparente y la masa total de los insectos. Para la obtención de los datos de calibración se construyó una gráfica representativa de los puntos; utilizando un beaker de 50 mililitros, donde se colocaron alícuotas crecientes de termitas muertas. El volumen (X) se determinó mediante el enrasado visual de la masa de insectos tras un ligero asentamiento mecánico para homogeneizar los espacios entre ellas. Inmediatamente después de registrar el volumen de cada alícuota, la masa total (Y) se cuantificó utilizando una balanza electrónica de precisión analítica con cuatro decimales (0.0001 g). Cada punto de la curva de regresión representa una medida, por lo tanto, el par ordenado de volumen medido y su masa correspondiente.
De forma complementaria, para calibrar la equivalencia en individuos, se determinó el peso promedio de una termita mediante 75 pesadas independientes de submuestras de 100 individuos cada una, distribuidas equitativamente entre los cinco tratamientos, sus cinco repeticiones y tres muestreos por termitero. Finalmente, la densidad poblacional absoluta para cualquier volumen evaluado en campo se estimó mediante la relación analítica entre la masa predicha por el modelo y el peso promedio por individuo.
Diseño del experimento y análisis estadístico. Se empleó un diseño completamente aleatorizado (DCA) en ambas fases del experimento. Los tratamientos corresponden a cuatro dosis de un bioinsumo y un testigo (aplicación de agua), con cinco repeticiones y la unidad experimental constituida por el termitero, retirado del árbol, y compuesto por cuatro fragmentos, dos de ellos muestreado en campo a los 25 días después de la aplicación del consorcio y los otros dos (en el laboratorio) a los 50 días después de la aplicación.
Las variables cuantificadas fueron: termitas vivas y termitas muertas por termitero, termitas muertas colonizadas, porcentaje de colonización y peso de 100 termitas. Los datos fueron analizados por estadística no paramétrica, por no presentar homocedasticidad tanto de varianza como de normalidad de los errores, empleando la prueba Kruskal-Wallis y estadística descriptiva. La comparación entre tratamientos se realizó mediante la prueba de comparación de rangos con 5 % de margen de error. Las poblaciones de termitas, en función de los tratamientos, fueron determinadas a través de la relación volumen-peso, mediante una ecuación de regresión lineal. Todos los procedimientos estadísticos fueron realizados utilizando el programa InfoStat versión 2020 (Di Rienzo et al., 2020).
RESULTADOS Y DISCUSIÓN
Fase de campo (25 días después de la aplicación)
Se presentaron diferencias significativas entre tratamientos para el número de termitas vivas, particularmente entre el testigo y la dosis más baja del consorcio (igual categoría estadística) y las dosis más altas del consorcio (Cuadro 1). Los tratamientos con dosis de 20 ml L-1 y 30 ml L-1 presentaron los menores conteos de termitas vivas, con mediana de 1 644 y 85 individuos respectivamente; siendo estadísticamente similares entre sí por la prueba de rangos y con la menor proporción de termitas vivas; mientras que, los tratamientos con la dosis de 5 ml L-1 y el testigo, presentaron las medianas con la mayor cantidad de termitas vivas.
| Tratamientos | Termitas vivas | Termitas muertas | |||
| H | p valor | H | p valor | ||
| 10.46 | 0.0328 | 1.62 | 0.4105 | ||
| Rangos medios | Mediana | Rangos medios | Mediana | ||
| 0 ml L-1 | 17.60 a | 29889.00 | 11.00 | 0.00 | |
| 5 ml L-1 | 19.20 a | 41003.00 | 16.00 | 0.00 | |
| 10 ml L-1 | 12.50 ab | 10629.00 | 11.00 | 0.00 | |
| 20 ml L-1 | 7.40 b | 1644.00 | 13.40 | 0.00 | |
| 30 ml L-1 | 8.30 b | 85.00 | 13.60 | 0.00 | |
La dosis de 10 ml L-1, presentó un comportamiento intermedio para ambos grupos estadísticos. También se observa la efectividad del consorcio de entomopatógenos (20 ml L-1 y 30 ml L-1) para reducir de manera significativa la población de termitas vivas en los termiteros en relación con el testigo y la dosis más baja del consorcio.
En el caso de las termitas muertas, no se registran diferencias significativas, debido a que los tratamientos según la prueba de Kruskal-Wallis, presentaron medianas con valor cero (Cuadro 1). El hecho de ocurrir disminución progresiva de las poblaciones de termitas, para la variable termitas vivas, en función del incremento de las dosis del biológico, indica que los consorcios con mayor dosis podrían tener un efecto irritante y/o repelente para el insecto.
La disminución observada en el número de termitas vivas, a los 25 días de la aplicación de los entomopatógenos ‒condiciones de campo‒, para las dosis de 10, 20 y 30 ml L-1, en relación con los tratamientos testigo ‒sin aplicación‒ y del biológico, en su dosis más baja, podría ser explicado, por la formación de compuestos orgánicos volátiles ‒COVs‒. Según Morath et al. (2012) se encuentran identificados más de 300 COVs de origen fúngico; entre estos compuestos se hallan grupos funcionales como cetonas, heterocíclicos, aldehídos, fenoles, ácidos, alcoholes, terpenos, tioalcoholes, tioesteres, compuestos aromáticos y ciclohexano, derivados de rutas metabólicas de la oxidación de la glucosa (Morath et al., 2012; Lemfack et al., 2013; Schmidt et al., 2015). Varias especies de hongos entomopatógenos son capaces de producir ácidos orgánicos y algunos de ellos han sido implicados en el proceso infectivo. Beauveria spp., Lecanicillium (Verticillium) lecanii, Isariafumosoroseus y Metarhizium anisopliae producen ácido oxálico (Hegedus y Khachatourians, 1995; Asaff et al., 2006). Otro compuesto importante detectado en algunos hongos entomopatógenos, entre ellos, Paecilomyces spp., y Metarhiziumanisopliae es el ácido 2.6-piridindicarboxilico, o ácido dipicolínico (Asaff et al., 2006).
Los compuestos orgánicos volátiles podrían ahuyentar las termitas, constituyendo un mecanismo potencial, por esta razón no se detectaron ejemplares muertos de la plaga en el interior de los termiteros; de modo similar, podría entenderse como un mecanismo de protección de sus nidos, salir y morir externamente para proteger la integridad de la colonia. Este evento, podría ser concebido como una evasión de las termitas al biológico; pero, es conveniente señalar que los termiteros fueron humedecidos, en consecuencia, las esporas de los entomopatógenos disueltos en la solución aplicada, estarían adheridos a su cutícula, lo que provoca la infección del insecto plaga, conduciéndolo a su muerte (Pucheta-Díaz et al., 2006), independiente del lugar hacia donde puedan trasladarse. Se ha señalado que las termitas han desarrollado estrategias de defensa para eludir o disminuir el impacto de los entomopatógenos (Hassan et al., 2024), tales como acicalamiento social, conductas de alarma, repelencia de las termitas hacia las esporas fúngicas, enterrar o evitar cadáveres infectados por hongos y respuestas inmunológicas, que desafían el control biológico contra las termitas.
Cristaldo y Toledo (2025) señalan que la mayoría de las termitas, específicamente las obreras y soldados, son ciegas o tienen una visión extremadamente deficiente, ya que viven y trabajan en la oscuridad. Dependen de otros sentidos, como el tacto, el olfato y las feromonas para comunicarse y buscar alimento. Solo las termitas reproductoras ‒reinas y reyes‒ desarrollan visión funcional. Los otros mecanismos de comunicación que les permite sobrevivir de manera eficiente en la naturaleza involucran a las feromonas, para enviar mensajes, dejar rastro, que les permite guiar a otras hacia la comida o para señalar peligro. También, se comunican tocándose sus antenas o cuerpos, y cuando son amenazadas, golpean la cabeza contra las paredes de sus túneles para generar vibraciones; de esta manera, advierten ante el peligro y huyen. Estas consideraciones fortalecen la propuesta de abandono de los termiteros por efecto de las concentraciones del biológico evaluado.
Fase de laboratorio (50 días después de la aplicación)
La comparación de rangos para las termitas vivas a los 50 días después de la aplicación (dda) mostró que los tratamientos con las dosis de 5, 10, 20 y 30 ml L-1, respectivamente del bioinsumo, tuvieron los menores valores de individuos vivos coincidiendo todos con medianas iguales a cero, a diferencia del tratamiento testigo, que presentó la mayor cantidad de termitas vivas.
| Tratamientos | 50 días después de la aplicación del biológico | |||||
| Termitas vivas | Termitas muertas | |||||
| H | P valor | H | P valor | |||
| 11.82 | 0.0003 | 10.19 | 0.0323 | |||
| Rangos medios | Mediana | Rangos medios | Mediana | |||
| 0 ml L-1 | 23.00 a | 16397.00 | 4.50 b | 0.00 | ||
| 5 ml L-1 | 10.00 b | 0.00 | 13.20 ab | 166.00 | ||
| 10 ml L-1 | 12.00 b | 0.00 | 13.10 ab | 121.00 | ||
| 20 ml L-1 | 10.00 b | 0.00 | 15.60 a | 400.00 | ||
| 30 ml L-1 | 10.00 b | 0.00 | 18.60 a | 550.00 | ||
En relación con las termitas muertas, se generaron diferencias significativas entre los tratamientos, y se constata que los tratamientos con 20 y 30 ml L-1, presentaron medianas de 400 y 550 individuos muertos, respectivamente e idénticos estadísticamente; mientras que, el tratamiento testigo, no presentó termitas muertas. Las dosis de 5 ml L-1 y 10 ml L-1, presentan valores de medianas de 166 y 121, respectivamente, con magnitudes intermedias para la prueba de rangos. La similitud estadística de las dosis más elevadas del bioinsumo, con los mayores valores de termitas muertas señalan la potencialidad de consorcio de entomopatógenos para causar la mayor proporción de mortalidad, como lo descrito por Amer et al. (2008).
Número de termitas muertas colonizadas y porcentaje de colonización
Se registran diferencias significativas para ambas variables (Cuadro 3).
| Tratamientos | Número de termitas muertas colonizadas | Porcentaje de colonización | |||
| H | p valor | H | p valor | ||
| 8.08 | 0.0441 | 7.78 | 0.0495 | ||
| Rangos medios | Mediana | Rangos medios | Mediana | ||
| 0 ml L-1 | --- | --- | --- | --- | |
| 5 ml L-1 | 5.40 b | 95.00 | 6.20 b | 75.00 | |
| 10 ml L-1 | 7.00 b | 180.00 | 7.25 b | 83.53 | |
| 20 ml L-1 | 10.50 ab | 400.00 | 9.00 ab | 83.33 | |
| 30 ml L-1 | 15.17 a | 1681.00 | 16.00 a | 100.00 | |
La dosis de 30 ml L-1 difiere estadísticamente de los tratamientos con 5 ml L-1 y 10 ml L-1 del consorcio tanto para termitas muertas colonizadas como en el porcentaje de colonización (mayor mediana y porcentaje de colonización). La dosis de 20 ml L-1 presenta un valor intermedio y con el tratamiento sin aplicación, no se registra evento de colonización (Cuadro 3).
En este momento (50 dda), es posible constatar la potencial ventaja que presenta el consorcio de entomopatógenos con dosis de 30 ml L-1, al verificar la cantidad de termitas muertas colonizadas por hongos del consorcio, así como el mayor porcentaje de colonización de las termitas. Ambas características indican las bondades del consorcio de entomopatógenos con cepas de Beauveria bassiana, Metarhizium anisopliae, Lecanicillium lecanii y Paecilomyces sp., para el control de termitas.
Los resultados obtenidos en esta investigación, en su fase de laboratorio, se consolidan con los obtenidos de manera individual para entomopatógenos, como los realizados in vitro para Beauveria bassiana (Zoberi y Grace, 1990) y Paecilomyces fumosoroseus (Chai, 1995) con altos niveles de patogenicidad sobre termitas; de manera similar, es relatado Metarhizium anisopliae, atacando termitas como Reticulitermes flavipes (Zoberi 1995) y Coptotermes formosanus (Sajap y Kaur, 1990) en condiciones naturales. En ese mismo sentido, Subekti et al. (2024) señalaron que las esporas de hongos entomopatógenos de Metarhizium anisopliae, Trichoderma harzianum y Beauveria bassiana, formuladas con nanocelulosa como ingrediente inerte, contribuyen a la liberación de los ingredientes activos como el ácido decanoico, ácido hexadecanoico, ácido palmítico y ácido octadecadienoico, para controlar termitas. Las concentraciones más altas asociadas con el uso de nanocelulosa aumentaron la mortalidad de termitas; donde, Metarhizium anisopliae, combinado con nanocelulosa al 60%, obtuvo el mejor porcentaje de control de termitas de Coptotermes curvignathus y Cryptotermescynocephalus, siendo que, para las condiciones de campo, Metarhiziumanisopliae fue el entomopatógeno que mostró mejor control de termitas, tanto subterráneas como en madera seca.
Resultados de Kin et al. (2017) describen a Metarhizium anisopliae causando entre 71 % y 84 % de infecciones en termitas a los 15 días después de la aplicación, en las condiciones de esta investigación los efectos sobre las poblaciones de termitas muertas colonizadas y porcentaje de colonización solo pudieron ser observados a los 50 días después de la aplicación del biológico en condiciones de campo, sobre muestras mantenidas en condiciones de laboratorio.
De manera similar, la patogenicidad de entomopatógenos sobre otros insectos plagas, ha sido señalada por Tavera et al. (2024), al evaluar la efectividad del hongo Beauveria bassiana, en el control de la broca del café, y explica que, en condiciones de laboratorio, se constató ser un eficiente controlador del insecto plaga; además, indicó que su eficiencia de control depende de la cepa utilizada, ya que con la más promisoria obtuvo 100 % de mortalidad, a los siete días después de inoculado el insecto plaga, además constató la formación de micelio en el 97.5 % de los individuos objeto de la investigación. Análogamente, Gómez (1999), realizó un trabajo de patogenicidad de entomopatógenos, señalando a Lecanicillium lecanii, como un hongo que infecta a una serie de insectos plagas, siendo también antagónico para algunas enfermedades como la roya. En el Perú, se aisló este hongo de diferentes plagas y cultivos en diversos lugares, las cepas más patogénicas y virulentas empleadas se usan para el control de moscas blancas, pulgones y trips. Este hongo se produce en forma masiva en diferentes laboratorios a nivel mundial, para su uso en manejo integrado de plagas, al ser compatible con otras medidas de control.
Gómez et al. (2014) señalan que los hongos entomopatógenos producen toxinas que matan al insecto; otros, que no poseen toxinas, matan al insecto al consumir todos sus nutrimentos. Las toxinas pueden ser enzimas ―lipasas, glicogenasas, amilasas y quitinasas―, o metabolitos secundarios, cuya producción es una propiedad genética de los hongos, pudiendo ser afectada por diferentes factores, secretadas en cantidades significativas tanto en el cuerpo del insecto como en medios de cultivo. La muerte del insecto generalmente ocurre antes de que el hongo colonice totalmente el hemocele (cavidad corporal principal), de esta manera, finaliza la fase parasítica e inicia la fase saprofítica.
Peso de 100 termitas (g)
No se registran diferencias para esta variable (Cuadro 4), lo que indica que el peso no varía en relación con las dosis del consorcio respecto al tratamiento sin aplicación.
| Tratamientos | Peso de 100 termitas (g) | ||||
| Rangos medios | C | H | p valor | ||
| 1.00 | 2.67 | 0.6140 | |||
| Mediana | P10 | P90 | |||
| 0 ml L-1 | 13.80 | 0.28 | 0.21 | 0.29 | |
| 5 ml L-1 | 8.80 | 0.19 | 0.13 | 0.30 | |
| 10 ml L-1 | 12.00 | 0.27 | 0.25 | 0.28 | |
| 20 ml L-1 | 12.25 | 0.26 | 0.24 | 0.49 | |
| 30 ml L-1 | 8.25 | 0.21 | 0.17 | 0.29 | |
| General | 0.25 | 0.17 | 0.29 | ||
Los resultados indican que las diferentes dosis del consorcio no provocaron cambios significativos en el peso de 100 termitas (H = 2.67; p = 0.6140). Aunque se observaron ligeras variaciones en los rangos medios y medianas entre tratamientos, no se evidencia un efecto del bioinsumo sobre esta variable. Los valores de mediana, así como los percentiles P10 y P90, indican una distribución relativamente homogénea de los pesos entre tratamientos, sugiriendo estabilidad fisiológica en las termitas evaluadas independientemente de la dosis aplicada.
La bibliografía consultada no registra eventos de discusión sobre esta variable; aunque, podría ser valiosa, en el sensu biológico, ya que este parámetro puede indicar la etapa en la que se encuentra el proceso de patogénesis del hongo en el interior del insecto plaga.
Relación volumen-peso de las termitas
Los valores obtenidos para la ecuación de regresión indican ajuste lineal positivo, con coeficiente de determinación (R2) igual a 85 % de acuerdo con el método de los mínimos cuadrados, con tasa de incremento de 0.59 por unidad de volumen, en relación con el peso (Figura 1), lo que respalda su uso como herramienta para estimar indirectamente la biomasa o abundancia de termitas a partir del volumen.
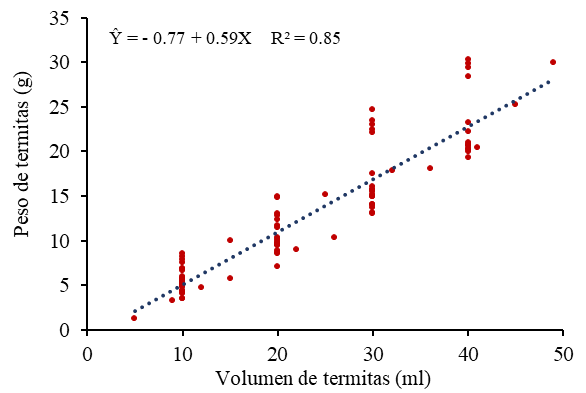
Figura 1.
Relación del peso de termita en función de su volumen.
La estrategia de control biológico con el uso de entomopatógenos es una técnica sustentable para el manejo de termitas, así lo afirma Awasthi et al. (2026) y Abebe (2002); además, Coêlho et al. (2023), plantean que afectaciones provocadas por temitas son reportadas de manera global, y que las investigaciones predominantes se corresponden con el uso de hongos entomopatógenos a nivel de laboratorio, con mayor frecuencia usando Metarhizium anisopliae y Beauveria bassiana.
CONCLUSIONES
La aplicación del consorcio de hongos entomopatógenos demostró diferente efectividad según el ambiente y tiempo de evaluación. A nivel de campo, las dosis de 20 ml L-1 y 30 ml L-1 ejercieron un posible efecto repelente a los 25 días después de la aplicación, que se evidencia por la evasión de las termitas, sin registrarse mortalidad ni colonización. En laboratorio, a los 50 días después de la aplicación, ambas dosis optimizaron el control, logrando la mayor cantidad de termitas muertas. La dosis de 30 ml L-1 destacó con los máximos porcentajes de colonización y mortalidad frente al tratamiento testigo, y se constituye como un bioinsecticida potencial para el manejo integrado de termitas.
REFERENCIAS BIBLIOGRÁFICAS
Abdu-Allah, G. M., Abou-Ghadir, N. M. F., Nasser, M. A. K. & Metwaly, M. R. (2015). Comparative efficiency of the fungi, Beauveria bassiana, Metarhizium anisopliae and the natural product spinosad, using three economic coleopterous stored grain insects. Egyptian Journal of Biological Pest Control, 25(3), 715-720. https://doi.org/10.13140/RG.2.1.3064.6161
Abebe, H. (2002). Potential of entomopathogenic fungi for the control of Macrotermes subhyalinus (Isoptera:Termitidae) [Tesis doctoral no publicada, Universität Hannover].
Acuña-Jiménez, M., Rosas-García, N., López-Meyer, M., Saínz-Hernández, J., Mundo-Ocampo, M., & García-Gutiérrez, C. (2015). Pathogenicity of microencapsulated insecticide from Beauveria bassiana and Metarhizium anisopliae against tobacco budworm, Heliothis virescens (Fabricius). Southwest Entomol., 40(3), 531-538.
Alcalá-Gómez, J., Cruz-Vázquez, C., Fernández-Ruvalcaba, M., Ángel-Sahagún, C., Vitela-Mendoza, I., & Ramos-Parra, M. (2017). Virulence of Metarhizium anisopliae and Beauveria bassiana isolates and the effects of fungal infection on the reproduction potential of Rhiphicephalus microplus engorged females. Biocontrol Sci. Technol., 27(8), 931-939.
Amer, M., El-Sayed, T., Bakheit, H., Moustafa, S., & El Sayed, Y. (2008). Pathogenicity and genetic variability of five entomopathogenic fungi against Spodoptera littoralis. Res. J. Agric. Biol. Sci., 4, 354-367.
Asaff, A., García-Rojas, C., Viniegra-González, G., & De la Torre, M. (2006). Carbon distribution and redirection of metabolism in P. fumosoroseus during solid-state and liquid fermentations. Process Biochem., 41, 1303-1310.
Awasthi, I., Gagneja, S., Patra, A., Capalash, N., & Prince, S. (2026). Biological Control Strategies for Termite Management. https://doi.org/10.20944/preprints202403.1842.v1
Barbosa, T., De Andrade, D., Polanczyk, R., & Duarte, R. (2018). Susceptibility of Tetranychus ogmophallos (Acari: Tetranychidae) to Beauveria bassiana and Metarhizium anisopliae. Fla Entomol., 101(2), 249-253. https://doi.org/10.1653/024.101.0215
Bidochka, M. J., Kasperski, J. E., & Wild, G. A. M. (1998). Occurrence of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana in soils from temperate and near-northern habitats. Canadian J. Bot., 76(7), 1198-1204. http://doi.org/10.1139/b98-115
Butt, T., Jackson, C., & Magan, N. (2001). Fungi as biocontrol agents, progress, problems and potential. CABI publishing.
Chai, Y. (1995). Estudios preliminares sobre la patogenicidad de algunos hongos entomógenos para Coptotermes formosanus. Revista China de Control Biológico, 11(2), 68-69.
Cristaldo, P. y Toledo, J. (2025). Ecología química de cupins. En A., Arab, I., Haifig, & T., Carrijo, (Ed.), Cupins da América do Sul (264-291). Editora UFABC. https://doi.org/10.5281/zenodo.15635957
Coêlho, C., Araujo, E., Brasil, L., Chagas, K., Silva, T., Orso, G., Mascarenhas, A., & Vendruscolo, J. (2023). Biological control of termites: a bibliometric and state-of-the-art review. Nativa Sinop, 11(1), 115-127. https://doi.org/10.31413/nativa.v11i1.14852
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., González, L. A., Tablada, M., & Robledo, C. W. (2020). InfoStat [Software]. Grupo InfoStat, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. http://www.infostat.com.ar
Espero, L. (1984). Survey of entomogenous fungi associated with termites. (Philippines). Scientific Journal, 6(1), 23-24.
Ferron, P. (1978). Biological control of insect pests by entomogenous fungi. Annual Review of Entomology, 23, 409-442. https://doi.org/10.1146/annurev.en.23.010178.002205
Fravel, D. (2005). Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol, 43, 337-359. https://doi.org/10.1146/annurev.phyto.43.032904.092924
Gómez, H. (1999). Aislamiento e identificación de hongos entomopatógenos de la “mosca blanca” Bemisia tabaci (Gennedius) (Homóptera: Aleyrodidae) en Lima. Rev. Per. Ent., 75, 516-525
Gómez, H., Zapata, A., Torres E. y Tenorio, M. (2014). Manual de producción y uso de hongos entomopatógenos. https://corporacionbiologica.info/wp-content/uploads/2020/11/Manual-de-Prod-y-Uso-de-Hong-Entomopa.pdf
Hassan, A., Li, Z., Zhou, X., Mo, J., & Huang, Q. (2024). Termite management by entomopathogenic fungi: Recent advances and future prospects. Current Research in Biotechnology, 7, 100-183.
Hegedus, D., & Khachatourians, G. (1995). The impact of biotechnology on hyphomycetous fungal insect biocontrol agents. Biotechnol. Adv., 13, 455-490.
Herrera, T. y Ulloa, M. (1990). El reino de los hongos, Micología básica y aplicada. Universidad Nacional Autónoma de México. http://repositorio.fciencias.unam.mx:8080/xmlui/handle/11154/149506
Khan, K., Jayaraj, B., & Gopalan, M. (1991). Mycopathogens for biological control of Odontotermes brunneus Hagen. Journal of Biological Control India, 5(1), 32-35. https://www.informaticsjournals.co.in/index.php/jbc/article/download/15268/13049/15913
Khan, K., Jayaraj, S., & Gopalan, M. (1990). Testing entomopathogenic fungi against the common agro-forestry termites. En G. K. Veeresh, B. Mallik, & C. A. Viraktamath (Eds.), Social insects and the environment (p. 636).
Kin, P., Moslim, R., Azmi, W., Kamarudin, N., & Ali, S. (2017). Genetic variation of entomopathogenic fungi, Metarhizium anisopliae and Isaria amoenerosea and their pathogenicity against subterranean termite, Coptotermes curvignathus. J. Oil Palm. Res., 29(1), 35-46.
Lemfack, M., Nickel, J., Dunkel, M., Preissner, R., & Piechulla, B. (2013). mVOC: a database of microbial volatiles. Nucleic acids research, 42(1), 744-748. https://doi.org/10.1093/nar/gkt1250
Li, H., Xu, Z., Deng,T., Chen, L., Li, J., Wei, J., & Mo, J. (2010). Species of termites (Isoptera) Attacking Trees in China. Sociobiology, 56(1), 109-120.
Lobo, L., Rodrigues, J., & Luz, C. (2016). Effectiveness of Metarhizium anisopliae formulations against dengue vectors under laboratory and field conditions. Bio. Sci. Technol., 26(3), 386-401.
Méndez, J., & Equihua, A. (2001). Diversidad y manejo de los termes de México (Hexápoda, Isóptera). Acta Zoológica Mexicana (Nueva Serie), Número especial 1, 173–187. https://doi.org/10.21829/azm.2001.8401850
Moino, A., & Alves, S. (1998). Efeito de imicacloprid e fipronil sobre Beauveria bassiana (Bals.) Vuill. E. Metarhizium anisopliae (Metsch.) Sorok. e no comportamento de limpeza de Heterotermes tenuis (Hagen). Anais da Sociedade Entomológica do Brasil, 27, 611-620.
Morath, S., Hung, R., & Bennett, J. (2012). Fungal volatile organic compounds: a review with emphasis on their biotechnological potential. Fungal Biology Reviews, 26(2-3), 73-83.
Morris, P. (2000). Integrated control of subterranean termites: The 6s approach group leader-Durability and Protection group. Forintek, Canada corporation.
Mugerwa, S. (2015). Magnitude of the termite problema and its potencial anthropogenic causes in Nakasongola distric of Uganda. Grassland Science, 61(2), 75-82.
Pérez-Levastida, M. (2014). Control de termitas con entomopatógenos. Universidad Central Marta Abreu de las Villas.
Petlamul, W., & Prasertsan, P. (2012). Evaluation of strains of Metarhizium anisopliae and Beauveria bassiana against Spodoptera litura on the basis of their virulence, germination rate, conidia production, radial growth and enzyme activity. Mycobiology, 40(2), 111-116. https://doi.org/10.5941/MYCO.2012.40.2.111
Pucheta-Díaz, M., Flores-Macías, A., Rodríguez-Navarro, S. y De la Torre, M. (2006). Mecanismo de acción de los hongos entomopatógenos. Interciencia Revista de ciencia y tecnología de América, 31(12), 856-860. https://www.redalyc.org/pdf/339/33901204.pdf
Qasim, M., Lin, Y., Fang, D., & Wang, L. (2015). Termites and microbial biological, control strategies. South Asia Journal of Multidisciplinary Studies, 1, 33-62.
Rath, A. (2000). The use of entomopathogenic fungi for control of termites. Biocontrol Sci. Technol., 10(5), 563-581.
Ricart, M., Bach, C. y Baltanás, R. (2015). Orden Isóptera. Revista IDE@-SEA, 49, 1-17. http://sea-entomologia.org/IDE@/revista_49.pdf
Rosales, C. (2001). Museo del Instituto de Zoología Agrícola Francisco Fernández Yépez. Universidad Central de Venezuela.
Rust, M., & Su, N. (2012). Managing social insects of urban importance. Annual Review of Entomology, 57, 355–375.
Rustiguel, C., Fernández-Bravo, M., Guimarães, L., & Moraga, E. (2018). Different strategies to kill the host presented by Metarhizium anisopliae and Beauveria bassiana. Can. J. Microbiol., 64(3), 191-200. https://doi.org/10.1139/cjm-2017-0517
Sajap, A. S. & Kaur, K. (1990). Histopathology of Metarhizium anisoplia, an entomopathogenic fungus, infection in-the Termite, Coptotermes curvignathus. Pertanika, 13(3), 331-334. https://psasir.upm.edu.my/id/eprint/2839/1/Histopathology_of_Metarhizium_anisopliae,_an_Entomopathogenic.pdf
Schmidt, R., Cordovez, V., De Boer, W., Raaijmakers, J., & Garbeva, P. (2015). Volatile affairs in microbial interactions. The ISME journal, 9(11), 2329-2335.
Subekti, N., Susilowati, A., Kusumaningrum, E., Fadhila, A., Salsabila, S., Zahra, C., Al Sabrina, N., Guswenrivo, I., Sanjaya, Y., Kurniawan, C., Iswanto, A., & Miranti, M. (2024). The application of entomopathogenic fungi Metarhizium anisopliae, Beauveria bassiana, and Trichoderma harzianum for Coptotermes curvignathus and Cryptotermes cynocephalus termite control in Indonesia. J. Korean Wood Sci. Technol., 52(3), 262-275. https://doi.org/10.5658/WOOD.2024.52.3.262
Tanada, Y., & Kaya, H. (1993). Insect pathology. Academic Press.
Tavera, R., López, L. y Agueda, C. (2024). Efecto de diferentes aislamientos del hongo Beauveria bassiana (bals) Vuill. sobre broca del café Hypothenemus hampei Ferrari (Coleoptera: Scolytidae). APF, 13(2), 59-68. https://sodiaf.org.do/apf/index.php/apf/article/view/181/157
Wang, C., & Powell, J. (2004). Cellulose bait improves the effectiveness of Metarhizium anisopliae as a microbial control of termites (Isoptera: Rhinotermitidae). Biol Control, 30(2), 523-529. https://archive.sebs.rutgers.edu/wp-content/uploads/9-Metarhizium-BioControl.pdf
Zoberi, M. H., & Grace, J. K. (1990). Isolation of the pathogen Beauveria bassiana from Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiología, 16(3), 289-296. https://www.ctahr.hawaii.edu/gracek/pdfs/039.pdf
Zoberi, M. H. (1995). Metarhizium anisopliae, a fungal pathogen of Reticulitermes flavipes (Isoptera: Rhinotermitidae). Mycologia, 87(3), 354-359. https://doi.org/10.1080/00275514.1995.12026539
Notas de autor
Información adicional
redalyc-journal-id: 7858